


SPIOはDCの交差提示と移動を促進し、陰イオン性SPIOはインターロイキン-1βに関連するナノアジュバント効果に影響を与えます
要約
超常磁性酸化鉄ナノ粒子(SPIO)は、樹状細胞(DC)にロードすることにより、さまざまなナノアジュバントの担体として使用するために合成および探索されています。私たちの研究では、均質で超常磁性のナノ粒子はDCによる内在化の影響を受けやすく、SPIOパルスDCは優れた生体適合性とオボアルブミン(OVA)交差提示の能力を示しました。ここで、SPIOをロードしたDCがinvitroでDCの成熟と移動を促進できることを発見しました。 それぞれ正電荷と負電荷を示す3-アミノプロピルトリメトキシシラン(APTS)とメソ-2,3-ジメルカプトコハク酸(DMSA)でコーティングされたSPIOを準備しました。 SPIOの表面電荷がDCの抗原交差提示に影響を与える可能性があるかどうかを調査することを目的とした。さらに、反対に帯電したSPIOで処理した後、インターロイキン-1β(IL-1β)の形成を調べて、ナノアジュバントのメカニズムを特定しました。結論として、我々の結果は、SPIOが生体適合性であり、二次リンパ節へのDCの移動を誘発する可能性があることを示唆している。 APTSでコーティングされたSPIO(SPIO / A + )は、抗原の交差提示とT細胞の活性化を促進する優れたアジュバント能を示し、DMSAでコーティングされたナノ粒子(SPIO / D - )を上回りました。 )。このプロセスは、IL-1βの分泌に関連している可能性があります。私たちの研究は、ナノアジュバントの予測的修飾に関する洞察を提供します。これは、DCワクチンの設計に役立ち、ヒト用ワクチンへの応用のための新しいアジュバントの作成につながる可能性があります。
背景
アジュバントは、臨床的および実験的使用に広く適用されており、免疫刺激剤、受動的デポー、または必要な免疫応答を促進することができる媒体のいずれかとして長い間考えられてきました[1、2]。ここ数十年で、ナノ粒子、微生物製品、エマルジョン、サイトカイン、ポリマー、リポソームなど、多くの多様なクラスの化合物がアジュバントとして使用されてきました[3,4,5]。免疫アジュバント(ナノアジュバント)としてのナノ粒子の進化は、免疫応答の拡大に基づく抗原送達の注目すべき領域を表しています。すべてのタイプのナノ粒子の中で、超常磁性酸化鉄ナノ粒子(SPIO)は、優れた生体適合性、適切な表面構造、および柔軟なリガンド結合を備えています[6]。これらの特性により、磁気共鳴画像法(MRI)、標的化ドラッグデリバリー、温熱療法など、さまざまな生物医学分野に適用できます[7、8]。
主要な専門抗原提示細胞(APC)である樹状細胞(DC)は、特定の抗原に対する一次応答の後に免疫記憶が生成される細胞媒介適応免疫において重要な役割を果たし、この記憶は応答の増強につながりますその抗原とのその後の遭遇に[9、10]。さらに、DCは、主要組織適合遺伝子複合体クラスI(MHC-I)、交差提示と呼ばれるプロセスによって外因性抗原の提示を促進し、細胞傷害性Tリンパ球(CTL)を活性化することができます[11]。交差提示を目的とした可溶性抗原は、受容体を介したエンドサイトーシスによって内在化され、プロテアソーム分解とペプチドローディングのために細胞質に移動します。従来のDCワクチンは、可溶性抗原の輸送が比較的不十分なため、中程度の免疫応答を引き起こします。したがって、ナノアジュバントは、多くの研究研究でDCにおける交差提示を強化するための可溶性抗原の担体として探索されてきました[12、13、14]。
コーティング材料は、水性SPIO懸濁液の安定化とその後の機能化において重要な役割を果たします[15]。以前の研究では、正に帯電したSPIOは、免疫応答を強化するDCの交差提示能力を促進することが示され、反対に帯電した対応物の効果を上回りました[16、17]。異なる電荷を帯びたSPIOがDCの抗原交差提示に影響を与えるメカニズムは明らかにされていません。自然免疫に関与する原型の炎症性サイトカインであるインターロイキン-1β(IL-1β)は、病原体関連分子パターン(PAMP)およびダメージ関連分子パターン(DAMP)を感知すると、DCなどの免疫細胞から分泌される可能性があります[ 18]。 IL-1βの産生は、インフラマソーム、特にNLRP3(NACHT、LRR、およびPYDドメインを含むタンパク質3)によって厳密に媒介されます。 NLRP3の活性化は、カスパーゼ-1の産生を誘導し、カスパーゼ-1は不活性なプロIL-1βを活性型のIL-1βに切断します[19]。シリカ、二重壁カーボンナノチューブ、二酸化チタンなどの特定の無機ナノ粒子は、インフラマソーム形成を誘発する可能性があります[20、21、22]。以前の研究では、マグネタイトナノ粒子の表面電荷とそれらの細胞効率との相関関係が報告されています[23]。ここでは、SPIOをロードしたDCの異なる表面電荷もIL-1βの分泌に影響を与える可能性があると推測し、これらの表面電荷とDCの交差提示機能との関係を調査することを目的としています。
メソッドとマテリアル
SPIOの準備
SPIOを準備するために、以前に報告されたように簡単な共沈法が採用されました[24]。簡単に言えば、FeCl 3 の混合溶液 およびFeSO 4 (モル比Fe 3+ :Fe 2+ =2:1)を窒素雰囲気下で調製し、37°Cで30分間激しく攪拌しました。 Fe 3 の黒色の沈殿物 O 4 ナノ粒子が形成され、すぐに磁気分離を使用して蒸留水で5回洗浄されました。 Fe 3 O 4 次に、蒸留水に3 mg / mLの濃度でpH3で分散させました。最終的に、懸濁液を95°Cで(空気で)通気し、茶色のSPIOを分離しました。
DMSAでコーティングされたSPIO(SPIO / D - )およびAPTSでコーティングされたSPIO(SPIO / A + )は、SPIOをメソ-2,3-ジメルカプトコハク酸(DMSA)および3-アミノプロピルトリメトキシシラン(APTS)でそれぞれコーティングすることによって調製されました。 SPIO / Dの場合 − 、モル比1:40のDMSAの水溶液を100mLのSPIO溶液に添加しました。連続的に攪拌しながら4時間反応させた後、SPIO / D - 50°Cで500rpmの速度で分離されました。 SPIO / A + の場合 、APTSをSPIO溶液に0.2:1のモル比で加え、5時間激しく攪拌しました。その後、沈殿物を永久磁石で溶解し、脱イオン水で洗浄し、超音波で処理した。得られた溶液を水で繰り返し洗浄し、SPIO / A + 最後に、真空下、37°Cで粉末に乾燥させました。
マウス
C57BL / 6マウスと強化緑色蛍光タンパク質(EGFP)トランスジェニックC57BL / 6マウスは、南京大学のモデル動物研究センターから購入し、南京大学中央研究所の特定病原体除去(SPF)条件で飼育しました。すべての動物実験は、中国の南京大学医学部の動物管理使用委員会によって承認されたプロトコルに従って実施されました。
細胞培養
マウスDCは、以前に記載されたように[25]マウス骨髄から生成されました。簡単に説明すると、8週齢のC57BL / 6マウスの骨髄単球を大腿骨と脛骨から分離しました。続いて、細胞をRPMI-1640(Gibco、Thermo Fisher Scientific、マサチューセッツ州ウォルサム、米国)で、10%胎児ウシ血清(FBS)、10 ng / mLのマウス顆粒球マクロファージコロニー刺激因子(GM-CSF)とともに培養しました。; Gibco、米国)、および1 ng / mLのマウスインターロイキン-4(IL-4、PeproTech、ロッキーヒル、ニュージャージー州、米国)。培地は2日ごとに新しい培地と交換しました。未成熟なDCは、通常6日目に収集されました。EGFP-DCは、上記の方法に従ってEGFPトランスジェニックC57BL / 6マウスに由来しました。
解凍した凍結ヒト末梢血単核細胞(PBMC)を、完全培地、つまりX-VIVOTM 15(Lonza Group Ltd.、Basel、Switzerland)=1:1で一晩静置しました。密度勾配遠心分離後、PBMCを培地に4時間懸濁しました。付着細胞からのヒトDCは、ヒトGM-CSF(100 ng / mL; R&D Systems、ミネソタ州ミネアポリス、米国)およびヒトIL-4(10 ng / mL; PeproTech、ロッキーヒル、 NJ、米国)。培地の半分は2日目と4日目に交換されました。未成熟なヒトDCは5日目に採取されました。サイトメガロウイルスエプスタインバーウイルスおよびインフルエンザウイルス(CEF)特異的T細胞は、MHCを含むCMX培地で培養された浮遊細胞に由来しました。 -CEFペプチド(20 ng / mL、Panatecs、ハイルブロン、ドイツ、PA-CEF-002)を約48時間制限しました。 CEF特異的T細胞は、CMX培地で1000 U / mLのヒトIL-2(Peprotech、ロッキーヒル、ニュージャージー州、米国、200–02)によって12日間増殖されました。 IL-2の場合、培地は3日ごとに新しい培地と交換されました。
特性評価
SPIOの形態とサイズは、透過型電子顕微鏡(TEM、Advanced Microscopy Techniques、マサチューセッツ州ダンバーズ)を使用して決定されました。 X線回折(XRD)パターンを使用して、触媒のスペクトルを特定しました。ゼータ電位アッセイも測定して、さまざまなポリマーでコーティングされたナノ粒子の表面電荷を決定しました。 SPIO / A + およびSPIO / D − ナノ粒子は、3〜8の範囲のpH値で調製されました。ゼータ電位測定は、ゼータサイザーNano ZS90電位アナライザー(Malvern、UK)を使用して行われました。ナノ粒子の磁気特性は、振動試料型磁力計(Lakeshore 7407)を使用して37°Cで分析されました。
CPRGアッセイ
B 3 Z T細胞株(CD8 + T細胞ハイブリドーマ)は、T細胞受容体がH-2Kb MHC-I分子の存在下でオーバルブミン(OVA)258–265エピトープに結合すると、LacZ遺伝子を発現する可能性があります。 DC(2×10 4 )およびOVA(100μg/ mL、Sigma-Aldrich)をSPIO、SPIO / A + で培養しました 、またはSPIO / D − 37°Cで。 6時間後、DCをB 3 と共培養しました。 Z(2×10 5 )一晩。クロロフェノールレッド-β-ガラクトシダーゼアッセイ(CPRG、Sigma-Aldrich、米国)を実施して、B 3 のβ-ガラクトシダーゼ産生を測定しました。 Zセル。このアッセイでは、595 nmでの光学密度(OD)は、DCの抗原交差提示能力を示しています。
細胞アポトーシスアッセイ
蛍光活性化セルソーティング(FCS)を使用して、DCの生存率に対するさまざまな濃度のSPIOの影響を調査しました。簡単に説明すると、未成熟DCをアネキシンVおよびPIでマークされたSPIO(Biouniquer、CHN)とインキュベートし、DCでのアネキシンVおよびPIの発現をFCSで調べました。
InVivoでの蛍光強度イメージング
インビボでのDCの移動を調査するために、TNF-α(60 ng /マウス)を両後肢のフットパッドに事前注入しました( n =8)。 24時間後、SPIOラベルのEGFP-DC(2×10 6 )40μLのリン酸緩衝生理食塩水(PBS)を、C57BL / 6マウスの左側のフットパッドに注入し、同じ数の非標識EGFP-DCを右側に注入しました。リンパ節に移動するEGFP-DCのレベルを調べるために、Maestroイメージングシステム(CRi、米国マサチューセッツ州ウォーバーン)を採用しました。解剖されたリンパ節は、484nmの励起フィルターと507nmの発光フィルターを使用して観察されました。次に、緑色蛍光を表示する蛍光画像を、Living Imageソフトウェア(v 2.50; Caliper Corporation、Newton、MA、USA)で分析しました。共焦点顕微鏡分析を使用して、免疫組織化学を決定した。凍結リンパ節を5μmの厚さの切片に切断して固定し、切片を緑色蛍光タンパク質(GFP)抗体(Invitrogen)とインキュベートし、二次抗体としてヤギAlexa Fluor 488 nm抗体(Invitrogen)を使用しました。共焦点レーザー走査型顕微鏡(Fluoview、Fv10i、Olympus)を使用してサンプルを観察しました。
インビトロでのヒトDC抗原クロスプレゼンテーションアッセイ
培養ヒトDC(2×10 4 )を、SPIO / A + が取り付けられた96ウェルの丸底プレートにシードしました。 およびSPIO / D − サイトメガロウイルス(CMV)pp65タンパク質(1μg/ mL、MACS)と組み合わせたナノ粒子(100μg/ mL)を追加し、CMV pp65タンパク質とCEFペプチド(1μg/ mL、Thinkペプチド)を別々にコントロールとして使用しました。 6時間後、CEF特異的T細胞(2×10 5 )が人間のDCに12時間追加されました。ブレフェルディンA(10μg/ ml、Sigma-Aldrich、米国)をさらに6時間培地に保持しました。刺激と活性化の後、CEF特異的T細胞を収集し、CD3、CD4、CD8、およびIFN-γ(Invitrogen、USA)に対するヒト抗体で染色しました。染色後、サンプルを特注のLSR IIフローサイトメーター(BD、フランクリンレイクス、ニュージャージー州、米国)で分析しました。収集されたデータは、FlowJoソフトウェア(Tree Star、米国オレゴン州アッシュランド)によって分析されました。
酵素結合免疫吸着測定法
マウスIL-1β酵素結合免疫吸着測定法(ELISA)Ready SET-Goキット(eBioscience、米国)およびヒトIL-1βELISAキット(Thermo Fisher Scientific、米国マサチューセッツ州ウォルサム)を使用してILの分泌を測定しました。 DCによる-1β。 100μL/ウェルのIFN-γでコーティングされたELISAプレート(Costar、USA)を使用して、4°Cで一晩抗体を捕捉し、ELISAバッファーでブロックしました。次に、サンプルと標準液をウェルに加え、37°Cで2時間インキュベートしました。ビオチン化IFN-γが検出されました。サンプルは、外径が450 nmに設定されたマイクロプレートリーダー(BioTek、米国)を使用して測定されました。
統計分析
データは、社会科学統計パッケージ(SPSS 13.0、シカゴ、イリノイ、米国)を使用して分析されました。結果は平均±SDとして提示され、対照群と試験群の差は一元配置分散分析、両側学生の t によって評価されました。 テスト、および分散分析の二重因子分析。 * P での違い <0.05は統計的に有意であると見なされました。
結果
SPIOの特性評価
合成されたSPIOの形態とサイズをTEMで観察しました。 TEM画像は、SPIOの平均サイズが8.7 nmで、球形であることを示しています(図1a)。 XRD分析では、標準のγ-Fe 2 と明確に一致する6つのピークが示されました。 O 3 反射(図1b)。振動磁力計の結果は、得られたSPIOが超常磁性の振る舞いを持ち、飽和磁化がFe 3 より60.4emu / g優れていることを示しました。 O 4 (図1c)。 DLSプロットは、SPIOのサイズ分布が溶液中で22 nmであることを示しました(図1d)。 DCにSPIOが含まれていることを確認するために、プルシアンブルー染色を実行して、DCに鉄が含まれていることを確認しました(図1e)。
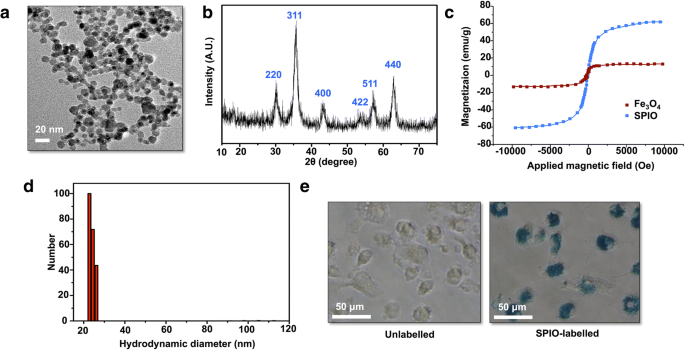
SPIOの特性評価。 a 得られたSPIOのTEM画像。 b 材料がγ-Fe 2 であることを示すナノ粒子触媒のXRDパターン O 3 。 c 得られたSPIOとFe 3 の磁化曲線 O 4 ナノ粒子。 d SPIOの流体力学的直径。 e 12時間のインキュベーション後の50μg/ mLSPIOで標識されたDCの形態:非標識DCおよびプルシアンブルー染色標識DC
SPIOがアクティブ化したDCのクロスプレゼンテーション
マウスシステムにおけるT細胞活性化に対するSPIO標識DCの効果をさらに研究するために、B 3 のレベル Z T細胞の活性化は、CPRGアッセイによってβ-ガラクトシダーゼの産生を調べることによって決定されました。この研究では、100μg/ mLのOVAの固定濃度と5つの用量比のSPIO(6時間後に1、5、10、25、および50μg/ mL)を採用しました。 SPIOの濃度が増加するにつれて、B 3 の活性化の程度 Z細胞は徐々に増加し、25μg/ mLで安定に達しました(図2a)。さまざまな濃度のSPIOで標識されたDCの生存率が影響を受けたかどうかを調べるために、DCおよびSPIOで標識されたDCを、アネキシンVおよびPI染色後にFCSを介して分析しました。結果は、10、25、および50μg/ mLのSPIO濃度でのアネキシンV / PI DCの合計パーセンテージは有意に異ならなかったが、アポトーシス細胞のパーセンテージは100μg/ mLSPIOをロードした後に増加したことを示しました(図2b )。さまざまな濃度のSPIOで標識されたDCの表面共刺激分子がFCSによって観察されました。 CD80およびCD86の発現は、SPIO標識なしのものと比較して、25μg/ mLで検出可能な増加を示しました(図2c)。 25μg/ mLSPIOで標識されたDCは、さまざまな時点で細胞アポトーシスの変化を示しませんでした(図2d)。以下の実験では25μg/ mLを使用しました。
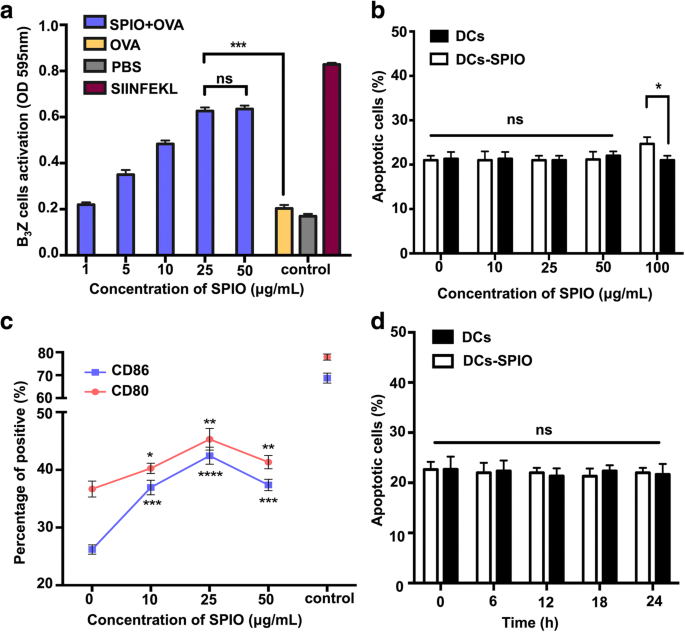
SPIOで標識した後のDC交差提示と生体適合性の影響。 a 異なるSPIO濃度で強化されたDCの交差提示。 b さまざまな濃度(10、25、50、および100μg/ mL)でFCSによって検査されたアポトーシスDCおよびSPIOラベル付きDC。 c CD11c + を含むDCの表現型 CD80 + およびCD11c + CD86 + SPIO(10、25、50μg / mL)で標識した後。 LPS(1μg/ mL)で刺激されたDCをポジティブコントロールとして使用しました。 d さまざまな時点(6、12、18、および24時間)でFCSによって検査された25μg/ mLのSPIOをロードしたアポトーシスDC
二次リンパ節でのSPIO強化EGFPシグナルによるEGFP-DCのラベル付け
EGFP-DCはinvitroでEGFPトランスジェニックマウスからの誘導に成功し、共焦点蛍光顕微鏡画像は、ほぼすべてのEGFP-DCが緑色蛍光を示したことを示しました(図3a)。 EGFP-DCの緑色蛍光がSPIOの影響を受けるかどうかを調べるために、FCSを実施しました。結果は、EGFP蛍光の発現がSPIO標識後に弱められなかったことを示しました(図3b、c)。次に、25μg/ mLのSPIO標識EGFP-DCをC57BL / 6マウスの右側の後足蹠に注入し、非標識EGFP-DCを反対側に注入しました。 EGFPシグナルは、それぞれセンチネルリンパ節と二次リンパ節である膝窩リンパ節(PLN)と鼠径リンパ節(ILN)で測定されました。結果は、センチネルリンパ節におけるSPIO標識EGFP-DCと非標識EGFP-DCの移動が7日目にピークに達したことを示しました。EGFPシグナルの有意差は2つのグループ間で観察されませんでしたが、有意な減少が検出されました。 ILNグループで検出されたEGFPシグナルは、4日目と7日目のSPIOラベル付きEGFP-DCグループのシグナルと一致しており、SPIOラベル付きDCが二次リンパ節に移動したことを示しています。ノード(図3d–f)。
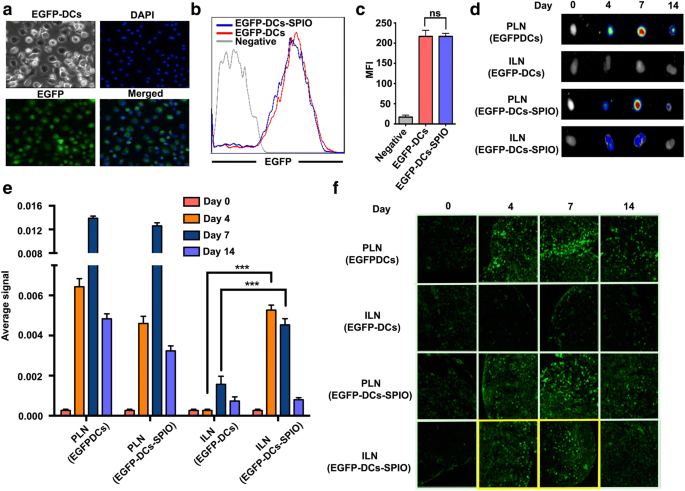
SPIOでラベル付けした後のEGFP-DCの移行と位置。 a 12時間のインキュベーション後、25μg/ mLのSPIOナノ粒子で標識されたEGFP-DCのレーザー走査型共焦点顕微鏡。 b 、 c 12時間後のSPIOラベル付きEGFP-DCの蛍光強度。異なる日にSPIO標識EGFP-DCを逆輸血した後のドレナージリンパ節のinvitro光学イメージング。 TNF-αは、EGFP-DCの移動を促進するために、事前にマウスの足の裏に最初に注射されました。 d – e SPIOの有無にかかわらずEGFP-DCの注射後の異なる日のリンパ節のinvitro光学イメージングおよび信号強度分析。 f 流入領域リンパ節のEGFP陽性細胞がレーザー共焦点顕微鏡で検出されました
異なる料金の変更されたSPIOの影響を受けるマウスDCのクロスプレゼンテーション
SPIOはAPTSまたはDMSAでコーティングされていました。 SPIOの表面電荷を確認するために、ゼータ電位特性を測定しました。 SPIO / A + のゼータ電位 およびSPIOは正でしたが、SPIO / D - のゼータ電位は正でした。 pH値が7のとき、溶液中で負の電荷を示しました(図4a)。異なる電荷のポリマーでコーティングされたSPIO標識DCの超微細構造をTEMで観察しました。 SPIOで処理されたDCは、未処理の細胞と比較して電子密度が高いように見え、多数のSPIOが細胞質に集まっていました。 SPIO / A + 細胞質内のエンドソームに飲み込まれたのに対し、SPIO / D - の量が多かった DCの細胞質で発見され、これらのナノ粒子のほぼすべてが、リソソームに似たいくつかの層状の膜構造に囲まれていました(図4b)。反対に帯電したSPIOが異なるレベルのIL-1βを誘発できるかどうかを調べました。結果によると、SPIO / A + がロードされたDC およびSPIO / D − IL-1βの用量依存的な分泌を誘発した。濃度に関係なく、SPIO / D - SPIO / A + よりも有意に高いレベルのIL-1βを誘導した (図4c)。 IL-1βとTLR3経路の間の明らかな相関関係により、T細胞活性化に対するIL-1βの効果をさらに研究するために、DCはTLR3ノックアウトマウスに由来し、SPIO / A + と共培養されました。 、SPIO / D − 、およびOVA。 SPIO / A + がロードされたDC + OVAはB 3 を効果的に活性化できます Z T細胞。これは、DC内のTLR3分子が交差提示に確実に必要であることを意味します(図4d)。
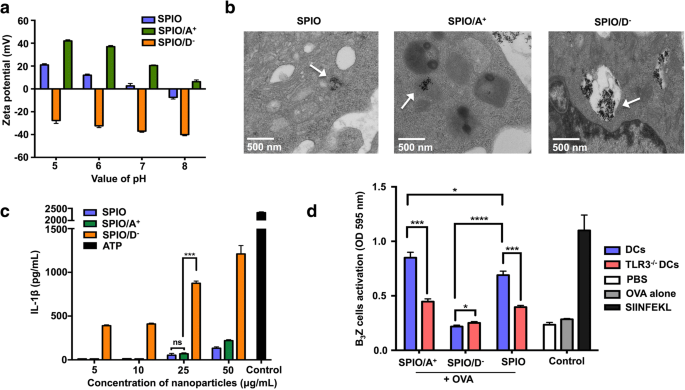
異なる電荷でコーティングされたSPIOは、DC交差提示とIL-1β分泌に影響を及ぼしました。 a 異なる電荷を持つ分子でコーティングされたSPIOのpH依存性ゼータ電位。 b TEM下のDCにおける異なる電荷を持つナノ粒子の位置。 c 異なる電荷を持つ分子でコーティングされたSPIOによって誘導されるIL-1β。 d SPIO / A + の交差提示 + OVAおよびSPIO / D − + TLR3経路を介したDCによるOVA
反対に帯電したコーティングで修飾されたSPIOは、ヒトDCによる抗原交差提示を促進し、IL-1βの影響を受けました
IL-1βと交差提示の関係を調べるために、カスパーゼ-1阻害剤YVADを選択し、50μMリポ多糖(LPS)で3時間前処理した後、ヒトDCのIL-1β分泌を有意に阻害できることを発見しました(図。5a)。さらに、YVADがSPIO / A + によって引き起こされるヒトDCからのIL-1βの分泌を阻害する可能性があることを発見しました。 およびSPIO / D − (図5b)。次に、ヒトDCを効果的なAPCとして使用し、CMVpp65をCEF特異的T細胞に交差提示できるかどうかを調査しました。抗原の交差提示を測定するために、細胞内染色(ICS)を導入して、抗原特異的CD3 + のパーセンテージを決定しました。 CD8 + IFN-γ + およびCD3 + CD4 + IFN-γ + FCSによるT細胞。 SPIO / A + がロードされたDC CMV pp65タンパク質と組み合わせると、より多くのCD3 + が誘導されました CD8 + IFN-γ + およびCD3 + CD4 + IFN-γ + SPIO / D - をロードしたDCよりもT細胞 。私たちのデータは、カスパーゼ-1阻害剤YVADがSPIO / D - によって誘導されるT細胞応答を有意に増加させることを示しました。 + CMV pp65タンパク質、SPIO / A + によって誘導されるT細胞応答を部分的に阻害した + CMV pp65タンパク質(図5c、d)。これらの結果は、効率的な交差提示には中程度のレベルのIL-1β活性化が必要であり、高レベルのIL-1β活性化がDCでの交差提示を抑制することを示しています。
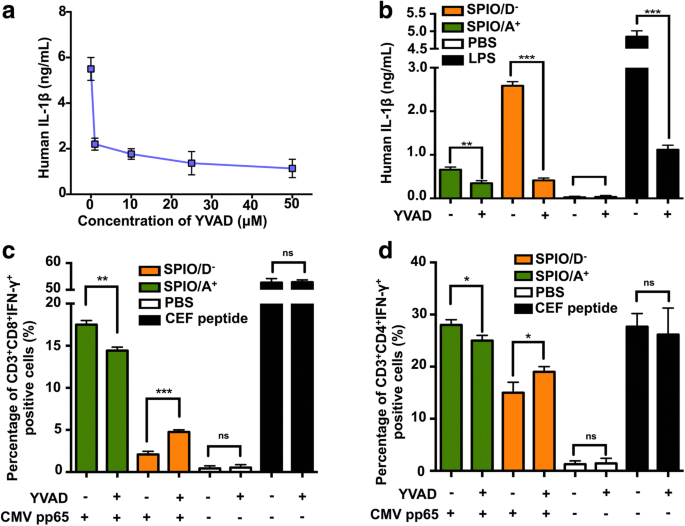
反対の電荷でコーティングされたSPIOは、IL-1β経路を介してヒトDCの機能に影響を与えます。 a LPSで3時間前処理した後、ヒトDCをYVADの濃度を上げながら(1、10、25、および50μM)インキュベートし、IL-1βのELISAのために上清を収集しました。 b YVADは、SPIO / D - を介してヒトDCからのIL-1βの分泌を阻害することができます 。 SPIO / A + 、およびSPIO / D − IL-1βの影響を受けるDC交差提示に影響を与えます。 c CD3 + CD8 + IFN-γ + および d CD3 + CD4 + IFN-γ + T細胞はFCSを使用した細胞内染色によって分析されました
ディスカッション
免疫療法は、バイオテクノロジーの開発以来、臨床および実験研究の研究の焦点となっています。しかし、免疫を誘発するように設計された従来のDCワクチンの以前の臨床試験では、十分な免疫応答が誘導されていません[26]。ナノ粒子は一種のアジュバントであり、DCの機能を促進してT細胞を活性化することができます[27]。私たちのSPIOの最も特徴的な機能には、生体適合性と、免疫アジュバントとしてのDCの標識の適用が含まれます。 TEM画像は、SPIOの平均サイズが乾燥状態で8.7 nmで、球形であることを示しています(図1a)。プルシアンブルー染色は、SPIOがDCによって貪食されやすいことを示し(図1e)、したがって摂取効率を示しています。
SPIOは、細胞標識、薬物送達、MRI、磁気温熱療法などの幅広い生物医学的用途があると報告されています[28]。これらはすべて生体適合性を必要とする用途です。したがって、SPIOで標識されたDCがDCアポトーシスに影響を与えるかどうかを調査する必要があります。私たちのデータは、DCが50μg/ mL未満のSPIOで標識された場合、DCアポトーシスに有意差がないことを示唆しました(図2b)。以下の研究では、モデルタンパク質としてOVAタンパク質を選択し、B 3 SPIOがDCの交差提示に影響を与えることができるかどうかを観察するための効果的なT細胞としてのZT細胞。私たちの結果は、SPIOでラベル付けされたDCがOVAの交差提示とB 3 の活性化を著しく促進できることを示しています ZTセル。成熟DCのマーカーである表面分子CD80およびCD86は、T細胞の活性化に必要な2つの重要な共刺激因子です。 25μg/ mLで、活性化されたB 3 Z T細胞は、ナノ粒子の投与量とともに増加し、最終的には安定しました(図2a)。さらに、DCの表面でのCD80およびCD86の発現は最大に達しました(図2c)。したがって、25μg/ mLの濃度を採用し、DCのアポトーシスは時間に依存しないことがわかり(図2d)、SPIOの優れたナノアジュバント機能と生体適合性が証明されました。
DC移行に対するSPIOの影響を調べるために、共焦点蛍光顕微鏡下で緑色蛍光を示すEGFP-DCを使用して、25μg/ mLのSPIOと12時間共培養しました。 SPIO標識後、EGFP蛍光の発現が弱まらないことを観察しました。最終的に、DCの二次リンパ器官への移動は、DCベースのワクチンの有効性を評価するための重要なパラメーターです。以前の研究では、最初の24時間で、SPIO標識DCは二次リンパ節で有意な量で検出されませんでした[29]。この現象が時間とともに変化するかどうかをさらに判断するために、ラベル付きとラベルなしの両方のEGFP-DCを収集し、マウスの足蹠に注入して、DCの移動におけるSPIOの可能性を評価しました。私たちの結果は、4日目と7日目に緑色蛍光がILNに現れたことを示し(図3d–f)、SPIOがEGFP-DCの二次リンパ節への移動を促進できることを示しています。この特性は、腫瘍の転移を制限し、より多くのリンパ節でより多くのCTL細胞を活性化するために適用できます。
私たちの研究では、反対に帯電したSPIOのアジュバント特性と、DCの機能の変化の根底にある考えられるメカニズムを調査しました。酸化鉄ナノ粒子の表面電荷は、細胞の取り込み効率に影響を与えることが示されています[30、31]。私たちの以前の研究では、カチオン性SPIOが抗原の交差提示を強化し、したがってT細胞の活性化を促進し、アニオン性SPIOがオートファジーと関連していることを報告しました。 [16]いくつかの金属ナノ粒子は炎症反応を誘発する可能性があります[32、33]。酸化鉄ナノ粒子は、NLRP3を活性化し、リソソームの膜を破壊し、カテプシンBを放出し、IL-1β分泌を誘導することが報告されています[34]。したがって、反対に帯電したSPIOナノ粒子は、マウスとヒトの両方のDCでIL-1βの産生を刺激する際に異なる機能を果たす可能性があると仮定しました。図4aでは、ゼータ電位分析により、APTSでコーティングされたSPIOが正の電荷を帯びているのに対し、DMSAでコーティングされたSPIOは負の電荷を帯びていることがわかりました。 TEMの下で、異なる電荷を持つSPIOが細胞質内の異なる位置に集まっていることがわかりました(図4b)。 SPIO / A + の交差提示の違いを調査するには およびSPIO / D − 、我々は、異なる電荷を帯びたナノ粒子によって誘発されたマウスDCにおけるIL-1β分泌を調査しました。 SPIO / A + による治療後のマウスDCの反応を評価しました およびSPIO / D − 。 SPIO / Dへの暴露 − SPIO / A + への曝露と比較して、IL-1β分泌の明白な活性化を誘導しました (図4c)。この現象は、SPIO / D - によって誘導される抗原の交差提示を部分的に明らかにしました。 SPIO / A + によって誘発されるほど効率的ではありません 。増加するB 3 のCPRG分析に加えて マウスDCとSPIO / A + の共培養後のZT細胞の活性化 + OVAは、正に帯電したナノ粒子が免疫アジュバントとして使用された場合に、より優れた性能を発揮できることを実証しました。 Toll様受容体(TLR)は、TLR3などの免疫応答を誘発するために重要であり、最終的にはインフラマソームの産生、カスパーゼ-1タンパク質の活性化、およびサイトカインIL-1βの分泌を引き起こす可能性があります[35]。 TLR3がナノアジュバントの影響を受ける抗原交差提示に関連しているかどうかを示すために、TLR3ノックアウトDCを研究に採用しました。 TLR3 -/- SPIO / A + がロードされたDC + OVAとSPIO + OVAはB 3 を効果的に刺激しました Z T細胞の活性化(図4d)。これは、DCのTLR3分子が交差提示に必要であることを示しています。まとめると、これらの結果は、活性化された免疫応答に対する相乗効果を調査する際に、ナノ粒子の表面にある異なる電荷のポリマーを考慮する必要があることを明らかにしました。
私たちのデータは、反対に帯電したナノ粒子がIL-1βの産生を誘発する可能性があることを示していますが、SPIO / D - IL-1βの分泌を過剰に活性化する可能性があります。ただし、SPIO / A + と比較して 、SPIO / D − より低いレベルのB 3 を誘発した ZT細胞の活性化。したがって、異常なIL-1β産生がDCの細胞機能障害に寄与する可能性があると推測します。ナノマテリアルアジュバントに関するほとんどの研究は、マウスからのDCを利用していますが、ヒトからのDCを使用したものはほとんどありません[36、37]。仮説をさらに検証するために、効果的なカスパーゼ-1阻害剤であることが証明されているYVADを使用して(図5a、b)、ヒトDCからのIL-1β分泌を抑制しました。モデル抗原としてCMVpp65タンパク質を選択し、invitroで増殖させたCEF特異的T細胞をレスポンダー細胞として使用しました。 CMV pp65タンパク質は、CD8 + を容易に活性化する免疫学的優勢タンパク質です。 およびCD4 + サイトカイン、特にIFN-γを産生するT細胞[38]。 CEFペプチドをポジティブコントロールとして使用し、YVADの添加がT細胞に直接提示できるため、T細胞の活性化に影響を与えないことを確認しました。興味深いことに、YVADを使用すると、SPIO / A + のT細胞応答 グループはわずかに抑制されましたが、SPIO / D - の応答は グループが増加しました(図5c、d)。 DC交差提示に対するIL-1βの影響は、SPIO / A + によって誘導される低レベルのIL-1βの決定的な証拠を提供しました。 抗原の交差提示に必要であり、細胞質ゾル中の高レベルのIL-1βはDCの機能を阻害します。
結論
図6の図の要約に示されているように、SPIOは、DCの成熟、移行、および交差提示を促進できます。中程度のIL-1β活性は、DCの抗原交差提示に部分的に関連しています。さらに、負に帯電したナノ粒子は過剰なIL-1βを活性化し、その後DCの機能を阻害する可能性があります。要約すると、我々の結果は、SPIOが多くの生物学的特性を示し、有望なアジュバントの可能性を持っていることを示しています。これらの調査結果は、将来のDCワクチンの開発に最適なナノアジュバントの選択を特定するのに役立ちます。
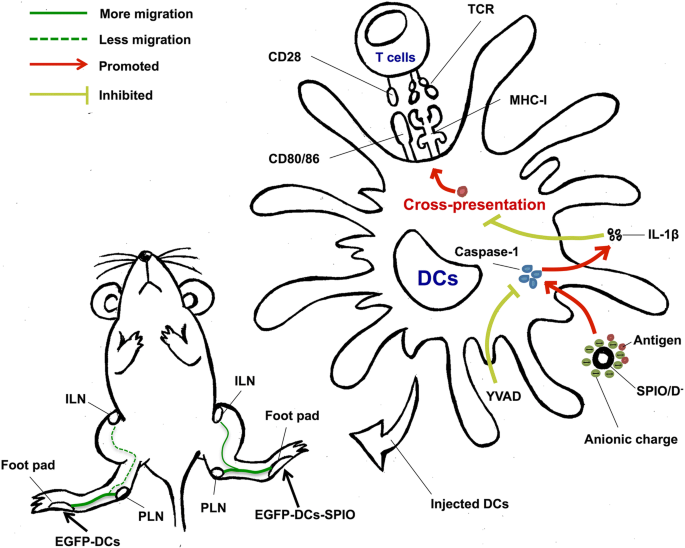
DCのナノアジュバントとしてのSPIOのグラフィカルな要約。 SPIOは、DCのロードを促進することにより、DCの機能を強化します。したがって、SPIOで標識されたDCは、免疫器官で活性のあるILNに移動することができ、癌免疫療法の新しいアプローチを提供する可能性があります。陰イオン性に帯電したSPIOは、DCでカスパーゼ-1をトリガーすることにより、保護的なIL-1β応答を活性化し、それによって活性T細胞への抗原提示を損ないます
略語
- APC:
-
抗原提示細胞
- APTS:
-
3-アミノプロピルトリメトキシシラン
- ATP:
-
アデノシン三リン酸
- CPRG:
-
クロロフェノールレッド-β-d-ガラクトピラノシド
- CTL:
-
細胞傷害性T細胞
- DAMP:
-
ダメージ関連分子パターン
- DC:
-
樹状細胞
- DMSA:
-
Meso-2,3-ジメルカプトコハク酸
- EGFP:
-
強化された緑色蛍光タンパク質
- IL-1β:
-
インターロイキン-1β
- MHC-I:
-
主要組織適合遺伝子複合体クラスI
- NLRP3:
-
NACHT、LRR、およびPYDドメインを含むタンパク質3
- OVA:
-
卵白アルブミン
- PAMP:
-
病原体関連分子パターン
- SPIO:
-
超常磁性酸化鉄ナノ粒子
- SPIO / A + :
-
APTSでコーティングされたSPIO
- SPIO / D − :
-
DMSAでコーティングされたSPIO
- TLR:
-
Toll様受容体
ナノマテリアル
- 定積分と微積分の基本定理
- IoTがインダストリー4.0をどのように変革するかと中小企業に対するIoTの影響
- Li-Nb-O化合物の調製と光触媒性能に及ぼすLi / Nb比の影響
- 微結晶およびナノセルロースの構造と誘電特性に及ぼす水の影響
- Al2O3 / ZnOナノラミネートの形態的、光学的、および電気的特性に及ぼす二重層の厚さの影響
- メタマテリアルにおける表面プラズモンポラリトンと磁気双極子共鳴の結合効果
- ポリオール媒介プロセスによるZnOナノクリップの製造と特性評価
- ポリマーナノコンポジットの界面/相間特性および引張強度に及ぼすナノ粒子のサイズおよび凝集/凝集の影響
- ペロブスカイト太陽電池の変換効率に及ぼすTiO2コンパクト層に埋め込まれた異なるサイズと濃度のAgナノ粒子の影響
- ドットとダッシュに別れを告げて、光ストレージメディアを強化する
- プラズマと酸素/燃料の燃焼効果