


家禽および他の単胃動物におけるNano-Seの同化と作用:腸内細菌叢は答えですか?
要約
最近、家畜と魚の栄養におけるnano-Seの役割に焦点を当てた包括的なレビュー論文がNanoscale ResearchLettersに掲載されました。著者らは、nano-Seの生産に関連する問題と、動物産業および医学におけるその可能な応用について詳細に説明しました。ただし、nano-Seの作用の分子メカニズムは説明されておらず、nano-Seがどのようにアクティブなセレノプロテインに変換されるかという問題は解決されていません。腸内細菌叢は、ナノSeを亜セレン酸塩H 2 に変換できる可能性が高いようです。 セレノプロテインの次の合成を伴うSeまたはSe-リン酸。この可能性はさらに詳細に研究する必要があり、動物/家禽/魚の栄養におけるSeの供給源としてのnano-Seの長所と短所は批判的な評価を待っています。
背景
最近、家畜と魚の栄養におけるnano-Seの役割に焦点を当てた包括的なレビュー論文がNanoscale Research Letters [1]に掲載されました。著者らは、nano-Seの生産に関連する問題と、動物産業および医学におけるその可能な応用について詳細に説明しました。確かに、ナノ粒子として提示される多くの分子は、そのような粒子の新しい特性のために異常な振る舞いをすることが十分に確立されています。実際、動物の栄養に関連する公開された研究のほとんどでは、ナノ粒子のサイズは100nm未満です。このような状態では、粒子のサイズが非常に小さいため、粒子は多くの生物学的障壁を貫通し、Ag、チタン、Seなどのさまざまな元素の供給システムとして使用できます。
この点で、Seは特に興味深いものです。これは、世界中のほとんどの地域でこの元素が不足していることが証明されているためです[2]。多くの場合、土壌中のSe含有量は大きく変動するため、欠乏は人間の創造物であり、さらに重要なことは、土壌からのSeの利用可能性がさらに変動することです。実際、農業慣行の結果としての土壌酸性化と硫黄含有肥料の使用は、植物のSeの利用可能性を大幅に低下させ、穀物(小麦、トウモロコシ、大麦)と油糧種子(大豆)が生息する家畜によるSe消費量の低下につながります。食事の大部分を占めています。そのため、家禽、家畜、魚の市販飼料のSe補給が一般的になり、1970年代から使用されています。確かに、ビタミンミネラルプレミックスは、卵、肉/魚、牛乳の商業生産のためのSeの主要な供給源です。
nano-Seの話に関連するのは、サプリメントに含まれるSeの供給源です。実際、Seの主な補足形態は、銅生産の副産物である亜セレン酸ナトリウムです。 40年以上の使用で、亜セレン酸ナトリウムはその長所と短所を明確に示しています。実際、市販の飼料Se補給により、臨床的欠乏の兆候を示す家畜のSe欠乏は事実上消失しました。例外は反芻動物で、亜セレン酸ナトリウムが酸性のルーメン環境で沈殿する可能性があります。また、飼料サプリメントの使用に関する技術的な制限が原因です。 Se欠乏症は今でも世界中で見られ、セレン注射やボーラスなどの用途が使用されています。ただし、前述のレビューで正しく言及されているように、Seは、特に生産性の高い家畜や家禽の現代の遺伝学にとって、商業的に関連するストレス条件において非常に重要です。
今日、動物産業は栄養不足の予防から、Seを含む重要な栄養素の動物の正確な要件を満たすことへと移行しています。確かに、「正確な栄養」はこの概念を説明する用語です。このような状況では、亜セレン酸ナトリウム(およびセレン酸塩)が飼料サプリメントとしてさまざまな欠点を持っていることが明らかになりました。まず第一に、それは非常に反応性の高い化合物であり、アスコルビン酸やいくつかの飼料成分を含むさまざまな栄養素によって、飼料/プレミックスで利用できない金属形態に還元することができます。また、飼料の水分に溶解し、揮発性化合物に変換されて失われる可能性があります。第二に、亜セレン酸ナトリウムは用量依存的に酸化促進特性を持っており、動物/ニワトリの腸に悪影響を与える可能性があります。最後に、亜セレン酸ナトリウムの形のセレンは、胎盤を介して胎児への移行が不十分であり、Seの必要量が増加するが、飼料消費量が通常減少するストレス状態で使用できるSeの蓄えを体内に構築することはできません。亜セレン酸ナトリウムは、動物や人間にも過剰に毒性があります。しかし、Seの毒性が動物産業にとって問題であるという前述のレビュー[1]の結論には同意しません。これは、計算またはフィードの混合における人為的エラーにのみ関連しています。セレンの安全なウィンドウは非常に狭いですが(家禽と豚のSe補給の通常の投与量は約0.3 ppmですが、亜セレン酸ナトリウムの投与量が1〜2 ppmを超えると悪影響が見られます)、最新の飼料工場設備では毒性の問題を回避する良好な飼料混合の機会。
分析科学の進歩は、主要な飼料成分中のSeの主な形態がSeMetであり、トウモロコシ、大豆、小麦、大麦などの全Seの50%以上を占めることを発見する原動力でした[2]。したがって、進化の過程で、動物の消化器系はこの形態のセレンに適応し、その結果、SeMetは食事からより効率的に吸収され、体のタンパク質に非特異的に組み込まれるとSeが蓄えられます。実際、Se-Yeast、純粋なSeMet、OH-SeMet(2-ヒドロキシ-4-(メチルチオ)ブタン酸-Se)など、さまざまな有機Se源が市場に登場し、OH-SeMetが最大の効果を示しました。
nano-Seの問題に関する知識の主な欠如は、その代謝、特にH 2 への変換に関連しています。 次のSeCys合成およびセレノプロテインへの取り込みを伴うSe。レビュー[1]では、グルタチオンペルオキシダーゼを含むセレノプロテインはごくわずかしか言及されていませんが、ヒトと動物では少なくとも25のセレノプロテインが同定されています。人間/動物の栄養におけるSeの主な役割は、独特の触媒特性を持つセレノプロテインの合成に関連していると一般に認められており、それらの半分以上がレドックスバランスの維持と抗酸化防御に関与しています[3]。前述のレビュー[1]では、nano-Seの直接的な抗酸化特性(ROSの還元)が、その作用の考えられるメカニズムとして言及されています。ただし、肝臓や筋肉を含む主要な動物/家禽組織のSe濃度は、通常、10μMの範囲の新鮮な組織800〜900 ng / gを超えず、血漿中のSeレベルは約0.2〜0.3μg /です。 mlまたは2〜3μM、in vitroでのnano-Seの抗酸化特性に焦点を当てた最も引用された研究では、Se濃度がテストされ、5〜10倍高い抗酸化効果を示しました[4]。さらに、抗酸化化合物が効果的なフリーラジカルスカベンジャーであるためには、適切な場所で適切なタイミングで適切な濃度の抗酸化剤を使用することが重要です。これは問題をさらに複雑にするため、nano-Seが生体系に直接AO効果をもたらす可能性は低いと思われます。
したがって、動物の食事で使用される他の形態のセレンと同様に、nano-Seの抗酸化作用はセレノプロテイン遺伝子の発現とタンパク質合成に関連しています。実際、レビュー[1]には、GSH-Px活性に対するnano-Seのプラスの効果を確認するいくつかの参考文献があります。ここで問題はどのように nano-Seは活性セレノプロテインに変換されます。レビュー[1]には、ナノSeを次のSeタンパク質合成でセレノホスフェートに変換できるという提案があります(参照なし)。この提案は実験的に証明されるべきです。 SeMetは人体/動物の体内で合成できないため、nano-SeをSeMetに変換できるという2番目の提案は根本的に間違っています。植物とバクテリアだけがそれを作り出すことができます[5]。
腸内細菌叢がナノSeを亜セレン酸塩/亜セレン酸塩に酸化したり、H 2 に還元したりできるという非常に魅力的な提案があります。 セレノプロテインの次の合成を伴うSe [6、7]。最近、ナノSe粒子が溶解し、吸収前に微生物叢の存在下で腸内でSeの無機オキソアニオンに酸化されることを証明するいくつかの実験的証拠が提供されました[6]。さらに、内因性セレンナノ粒子(SeNP)の細胞内動的サイクルを示す架空の概略図が提案されました[7]。実際、元素セレンはスーパーオキシドラジカルによって亜セレン酸塩に再酸化される可能性があることが示唆されました。腸内細菌叢がこのプロセスを実行/加速できる可能性もあります。実際、Seの4つの主要な生物学的変換は、還元、酸化、メチル化、脱メチル化など、自然界で発生することが証明されています[8]。過去30年間に蓄積されたデータは、微生物が酸化反応と還元反応の両方に関与することにより、環境中のセレンサイクルで主要な役割を果たしていることを明確に示しています[9]。興味深いことに、Se 0 の微生物による酸化 Se 4+ へ 未確認の独立栄養細菌のグループによって、90年以上前に発見されました[10]。さらに、従属栄養細菌 Bacillus megaterium による元素セレンの亜セレン酸塩への酸化 、土壌から分離されたものはずっと後に報告されました[11]。確かに、Se 0 土壌中の酸化は比較的遅い速度で起こり、本質的に大部分が生物的であり、両方のSeO 3 を生成することが示されました。 2- およびSeO 4 2- [12]。さらに、元素セレン(Se 0 )の微生物による酸化 )化学ヘテロトロフおよび化学合成独立栄養性チオバチルスにより、 75 を使用して確認されました Se 0 トレーサーとして[13]。著者らは、土壌スラリーがSe 0 を酸化できることを示しました。 SeO 3 を使用 2- およびSeO 4 2- 形成。興味深いことに、オートクレーブまたは化学処理による土壌中の微生物の不活化は、プロセスを阻害しました。さらに、硫黄酸化細菌 Thiobacillus の培養 ASN-1は、還元硫黄化合物からエネルギーを生成するために使用される酵素でSe(0)の酸化を実行することが示されています[13]。さらに、Mn(II)またはFe(II)を酸化できる従属栄養細菌(レプトスリックス MnB1)はSe 0 を酸化することが示されました SeO 3 の形成を伴う 2- 反応の主な生成物として。興味深いことに、反応は酢酸塩やブドウ糖などの電子供与体に依存することが示されました[13]。 Se 0 の酸化 さまざまなバクテリアによるものは完全には対処されておらず、重要な研究機会の領域です。実際、腸内に存在する多種多様な微生物は、さまざまなSe変換に必要な条件を提供します。たとえば、nano-Se粒子を乳酸菌( Lactobacillus delbrueckii )とインキュベートした場合 subsp 。ブルガリクス LB-12)、有機Se化合物(主にSeCysとSeMet)が生成され、ナノSe粒子も部分的に溶解し、おそらく細菌の細胞壁から排出される物質の助けを借りて、非代謝的に無機セレンに変換されました[14]。
あるいは、元素セレンは、亜セレン酸塩呼吸細菌(たとえば、 Bacillus selenitireducens )によって還元/セレン化物に変換することができます。 )。以下のように、電子供与体乳酸の酢酸塩への不完全な酸化を伴う細菌によって行われた前述の反応は、以下のように提示された[7]:
$$ {\ mathrm {C}} _ 2 {\ mathrm {H}} _ 4 {\ mathrm {OHCOO}} ^ {-} + 2 {\ mathrm {Se}} ^ 0 + 2 {\ mathrm {H}} _ 2 \ mathrm {O} \ to {\ mathrm {C} \ mathrm {H}} _3 {\ mathrm {C} \ mathrm {OO}} ^ {-} + 2 {\ mathrm {H} \ mathrm {Se}} ^ {-} + {{\ mathrm {H} \ mathrm {CO}} _ 3} ^ {-} + 3 {\ mathrm {H}} ^ {+} $$したがって、反応の自由エネルギー(∆G I )は-2.8 kcal / mol e - であることが示されています 。これは、 Bacillus selenitireducens で 、還元メカニズムには、Se特異的異化酵素を使用することによるエネルギー節約が含まれます[15]。確かに、亜セレン酸塩を呼吸する細菌、 Bacillus selenitireducens 、かなりのレベルのSe − 2 を生成する可能性があります (水性HSe-として)Se 0 を使用 基質として[16]。以前に、赤いセレンが義務的な好酸性菌 Thiobacillus ferrooxidans によって還元されることが実証されました。 酸性(pH 3)、嫌気性条件下でH 2 0.03μmol/ mgタンパク質/時間の速度でのSe生成[17]。さらに、別の嫌気性細菌 Veillonella atypica セレンのオキシアニオンを還元して元素セレンを形成し、バクテリアによってさらに還元されて反応性セレン化物を形成できることが示されました[18]。興味深いことに、45年以上前に、Micrococcus lactilyticus( Veillonella alcalescens )の抽出物が報告されました。 )コロイド状セレンをH 2 に定量的に還元することができました Se [19]。最近、YedEおよびYedFタンパク質をコードする遺伝子が、細菌を含む原核生物のSe代謝に関与する新しい候補遺伝子と見なされています[20]。実際、予測されるSeトランスポーターであるYedEとレドックスタンパク質であるYedFの両方が、細菌細胞におけるセレンの代謝変換に関与している可能性があります。したがって、元素セレンを酸化または還元するさまざまな微生物の主要な能力が証明されました(表1)。しかし、そのような反応が動物の腸で起こるかどうかという質問に答えるために、さらなる調査が必要です。興味深いことに、表1に記載されているすべての細菌種から、ベイロネラ属の嫌気性グラム陰性菌が 特別な注意が必要です。 確かに、ベイロネラ 温血動物の消化管に見られます[21]。実際、食用動物では、ベイロネラ 菌株は、ニワトリの上部消化管[23]および盲腸[24]を含む、消化管のすべてのセクション([22]およびその参考文献)の先住民として定期的に検出されます。確かに、動物の胃腸管におけるナノSeの可能な変換の運命とメカニズムを解明するために追加の研究が必要です。ナノSe代謝は、ナノ粒子コーティング剤を含むナノ粒子組成に依存します。セレノプロテイン合成へのナノSeの関与の提案されたスキームを図1に示します。吸収、分布、クリアランスなどのナノSe代謝の基本的な理解は、動物/家禽科学において非常に重要です[25]。
<図>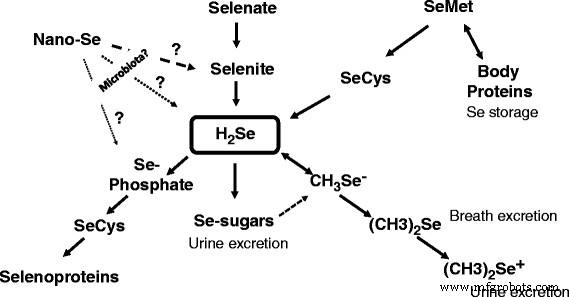
動物におけるさまざまな形態のSeの代謝変換を示す概略モデル。腸内細菌叢がナノSeを亜セレン酸塩H 2 に変換する可能性があることが示唆されています 次のSeCys合成およびセレノプロテインへの取り込みを伴うSeまたはSe-リン酸
哺乳類/鳥類の胃腸管には、総称して微生物叢として知られている何兆もの共生微生物が生息しています[26]。たとえば、鶏の腸管は十二指腸、空腸、回腸、盲腸、結腸で構成されており、前述の腸内細菌叢の濃度と組成には大きな違いがあります[27]。興味深いことに、盲腸は、門ファーミキューテスが優勢な最も複雑な微生物群集を特徴としています。 、バクテロイデス門 、放線菌 およびプロテオバクテリア [28]。一方、属レベルでは、すべての腸セクションにわたる主要な微生物属は乳酸桿菌であることが示されました。 、エンテロコッカス 、バクテロイデス 、およびコリネバクテリウム [27]。さらに、バクテロイデス Lactobacillus が盲腸で優勢なグループであることが示されましたが、 小腸セクション(十二指腸、空腸、回腸; [27])で優勢でした。この複雑な腸の生態系には、ナノSe粒子の酸化/還元を促進できるさまざまな微生物が存在する可能性があり、この仮定はさらなる調査を待っています。
一方の側から、いくつかの微生物は、微生物のセレノプロテインを合成し、この要素について宿主と競合するために、彼ら自身の必要性のためにSeを使用するでしょう。 Seは、ほとんどすべての細菌門のさまざまな生物にとって重要な元素であることはよく知られています。しかし、特徴づけられた細菌の3分の1だけが代謝にこの要素を使用しているようです[29]。実際、Se欠乏症では、利用可能なセレンをめぐって細菌と宿主の間で競争があり、無菌動物は従来のコロニーを形成した動物よりもセレンの必要量が少なくなります[30]。最近の結果は、食事によるSe補給が、既存の微生物叢の組成と多様性、および胃腸内細菌叢の確立の両方に影響を与える可能性があることを示しています[31]。たとえば、セレンが豊富なアルファルファ乾草を与えられた離乳した肉用牛は、対照動物と比較して、鼻の微生物相が豊富であることが示されました[32]。腸内細菌叢に対するさまざまな形態および濃度のSeの考えられる影響は、さらなる調査を待っています。
一方、微生物によるSeの能動的取り込みがあるようであり、このプロセスは使用されるSeの形態に依存します。実際、結腸の微生物叢によるSeMetの取り込みは、セレン酸の取り込みと比較してはるかに効率的であることが示されました[33]。ただし、細菌の組成と濃度が腸内のセレン化合物の吸収に影響を与えるかどうかは調査されていません。腸内細菌叢は、そのメチル化と元素のSe形成によって過剰なセレンの排泄に関与していると一般に認められています[34]。興味深いことに、SeMetを与えられたラットでは、この形態のSeが腸のすべての部分で見つかりました。しかし、回腸、盲腸、および結腸のSeのレベルは、プロバイオティクス治療後のラットの対応するセクションのレベルよりも有意に高かった[34]。これは、ストレプトコッカスサリバリウスを意味する可能性があります 、 Lactobacillus rhamnosus 、ラクトバチルスアシドフィルス および Bifidobacteriumlactis 主に遠方の腸でプロバイオティクスの影響を受けたSe代謝を伴って腸に送達された。実際、Seの代謝と同化における腸の各セグメントの役割を理解するには、Seの変換におけるさまざまな細菌の影響を特定する必要があります。さらに、大腸から小腸への上部からの液体の逆流が、H 2 の吸収の原因である可能性があります。 腸からのSeおよび他の形態のSe。実際、最近、鶏は総排泄腔から砂嚢にマーカーを移動させる可能性のある逆蠕動収縮を特徴とすることが示されました[35]。
細胞培養ではナノSeがセレノプロテイン(GSH-PxおよびTR活性)を増加させたため、セレノプロテイン合成へのナノSeの直接的な関与が期待できます。ただし、セレノプロテイン発現の優先順位の最近の理解は、この問題を複雑にする可能性があります。実際、多くのセレノプロテインは酸化ストレスによって調節されています。特に、GSH-Px1、GSH-Px4、およびTR1は、酸化ストレスに応答してアップレギュレーションされることが示され[36]、このような応答は、Seの供給が制限されている場合により顕著でした。前述の反応は、軽度の酸化ストレスにも当てはまるため、酸化ストレスのレベルにも依存しますが、非常に高いレベルでは、他のいくつかのメカニズムがアクティブになります[2]。
細胞培養では、そのようなセレノプロテインの発現/活性の増加は、ナノSeによって生成される酸化ストレスへの応答である可能性がありますが、Se供給の改善を反映するものではありません。したがって、細胞培養研究に基づく結果の解釈には注意が必要です。ナノSe補給による生体系のストレス関連の変化の追加の確認は、生体ナノSeが核因子(赤血球由来-2)様2(Nrf2)を活性化し、用量依存的および時間依存的に抗酸化合成に関与するその下流遺伝子の発現[37]。さらに、著者らは、Nrf2のノックダウンがそのようなナノSe粒子の抗酸化効果を大幅にブロックすることを発見しました。
家禽、家畜および魚の生産および繁殖成績に対するナノSe補給のプラスの効果を批判的に分析する場合、ほとんどの場合、Seは成績を改善せず、むしろ環境または栄養上の制約。多くの場合、動物実験は非常に低いSeバックグラウンドレベルで行われたため、利用可能な任意の形式でSeを追加すると、プラスの効果が得られます。
飼料添加物として使用されるnano-Seの商業化の課題は、次のとおりです。
- 1。
ナノSeの吸収、同化、および細胞、細胞内、遺伝子レベルでの作用の分子メカニズムを理解する必要があります。そのようなデータがなければ、再現性のある結果を得て、観察された効果の適切な説明を見つけることは困難です。たとえば、最近発表された論文では、ニワトリのナノSeの過剰摂取により、肝臓で影響を受けた遺伝子はごくわずか(18のタンパク質であり、Se代謝に直接関係するものはありません)です[38]。 SeMetを含む食事性Seの形態は、数百の遺伝子に影響を与える可能性があります。
- 2。
nano-SeがSeMetのように体内にSeの蓄えを構築できるかどうか、そしてそれらの蓄えがストレス状態で利用できるかどうかを理解することが重要です。
- 3。
同じサイズ、安定性、および適度に良好な(少なくとも6〜12か月)保存性のnano-Se粒子を提供できる技術を設計する必要があります[2]。
- 4。
微生物プロバイオティクスは、飼料添加物として使用されるナノセレンの変換に役立つ可能性があります。一方では、そのようなプロバイオティクスは、腸内でのナノSeの同化を助ける特定の微生物を含む可能性があります(例:ベイロネラ 種族)。一方、Seが豊富なプロバイオティクスは別の可能性がある可能性があります[34、39]
- 5。
副作用、リスク、環境問題に完全に対処する必要があります。実際、政策立案者や規制機関にnano-Seのナノ毒性の可能性について知らせるには、さらなる研究が必要です[40]。特に、乾燥した形の粒子のサイズが非常に小さいと、製品は非常にほこりっぽくなり、体内に入ると異常なnano-Se粒子の挙動が警告を発し、飼料工場の労働者の保護が主な懸念事項になります。腸内細菌叢は、メチル化と元素Se形成による過剰なセレンの排泄に関与しているため[34]、微生物叢の強化/調節は、ナノSe毒性の可能性に対処するための新たな地平を開く可能性があります。
- 6。
動物の栄養におけるnano-Seのプラスの効果は、その使用によって起こりうる有害な結果を覆い隠してはなりません。実際、さまざまな条件でのナノ粒子の挙動は大幅に異なる可能性があり、その挙動を制御する方法を理解する前に、幅広い産業規模でのナノ粒子の使用は不可能であるはずです。たとえば、ナノ粒子の吸収を検討する場合、ナノ粒子が腸内の他の物質の浸透を促進する特性を持っている可能性がある場合、いわゆるトロイの木馬効果に言及する必要があります[41]。飼料にはさまざまな「不要な」化合物が含まれており、腸はそれらの吸収を防ぐため、これはいくつかの問題を引き起こす可能性があります。確かに、腸内のnano-Seの動作は、さらに調査する必要があります。
- 7。
前述のレビュー[1]では、nano-Seの抗菌性と抗癌性にかなりの注意が払われており、ナノ粒子の独自の特性がさまざまな病状との闘いに役立つ可能性があります。確かに、nano-Seは、癌治療を含むさまざまな病状で使用される新薬と見なすことができますが、効果的な飼料添加物としての使用にはかなり疑問があります。今後の研究では、これらの重要な質問や懸念に答える必要があります。
結論
過去数年間、nano-Seに関するさまざまな論文が発表され、このトピックは急速に発展しています。ただし、この形態のSeが家禽/動物の商業生産への道を見つける前に、nano-Seがどのように活性セレノプロテインに変換されるかを理解して説明する必要があります。ナノSe作用の可能なメカニズム/経路の1つは、ナノSeを亜セレン酸塩H 2 に変換する可能性のある腸内細菌叢によって媒介される可能性があります。 SeまたはSe-リン酸と次のセレノプロテインの合成(図1)。自然界では、バクテリアが金属Se(Se 0 )を還元または酸化する可能性があるという証拠がいくつかあります。 )Se −2 の生成 またはSe +4 およびSe +6 それぞれ。セレドックスの変化に関与する微生物の中で、ベイロネラ属 鶏肉を含む食用動物の腸内にそのような細菌が存在するため、特別な注意を払う必要があります。腸内細菌がナノSeの同化と代謝に関与する可能性については、さらに詳細に調査する必要があり、動物/家禽の栄養におけるSeの供給源としてのナノSeの長所と短所は、さらに重要な評価を待っています。
ナノマテリアル
- DevOpsとクラウドがお互いを必要とする3つの理由。 DevCloudOps
- PVDFネジ、六角ボルト、六角ナット、その他のファスナーおよびカスタムパーツ
- 生物学的繁殖によって得られた動植物は、EPOで特許を取得できなくなりました
- メーカーや他のロボットユーザーにとってのロボットシミュレーションの3つの利点
- SAPS /4HANA®のプロセス自動化–希望、推進力、行動
- NexusIntegraと他のIoTおよびビッグデータプラットフォームとの違い
- PCB エンジニアが使用するソフトウェアやその他のツールは?
- ニッケル 718 組成とその他の主要な合金
- アルミニウム金属の残骸、ドロップ、およびその他の金属短絡を使用する利点
- トラック運送の安全上の懸念事項とその回避方法
- 横型マシニング センター (HMC) – 価格、コスト、およびその他の考慮事項