


HT29およびSPEV細胞株に対するAuナノ粒子の影響のinvitro研究
要約
細胞培養モデルは、ナノ粒子の潜在的な毒性および癌研究における基本的な調査のための優れたツールです。したがって、臨床現場でナノマテリアルを使用するには、AuNPの潜在的な毒性と人の健康への影響に関する情報が必要です。私たちの研究の目的は、上皮由来の細胞株に対するAuNPの影響を調べることです:連続的および発癌性。胚性ブタ腎臓上皮接種(SPEV)細胞株および結腸直腸癌細胞株(HT29)を使用した。試験培養では、細胞増殖、壊死/アポトーシス、および多細胞スフェロイドの生成が評価されました。 6〜12μg / mlのAuNP濃度は、SPEVおよびHT29細胞の増殖を減少させ、アポトーシスおよび壊死の初期および後期段階で細胞数を増加させることを実証しました。低濃度のAuNP(1〜3μg / ml)は、HT29およびSPEV細胞による多細胞スフェロイド形成を刺激することが示されました。ただし、AuNP濃度が高い(6〜12μg / ml)と、懸濁液中の細胞に細胞毒性と抗凝集効果の両方がありました。 AuNPの作用に対する大きな感度は、SPEV細胞(12μg/ ml)と比較してHT29(6μg/ ml)の系統によって示されました。 SPEVおよびHT29細胞株に対するAuNPの効果に関するこの実験的研究は、AuNPを介した抗がん治療へのさらなる応用を正当化するでしょう。
背景
金ナノ粒子(AuNP)の製造と調査は、金の幅広い治療への応用の可能性が高いだけでなく[1、2]、標的療法などの特定の生物医学的応用にも適しています[3、4]。最近の報告によると、AuNPの使用は、耐性発現のリスクを低減した新しい抗腫瘍療法の機会を提供します。したがって、いくつかの研究により、乳がん、肝臓がん、胃がん、結腸がん、肺がんに対するナノ粒子の抗腫瘍活性が証明されています[5、6]。
ナノ粒子(NP)は、細胞の運命を調節し、突然変異を誘発または防止し、細胞間コミュニケーションを開始し、細胞構造を調節できることが知られています[7、8]。さらに、AuNPは、生体適合性と抗腫瘍活性により、他の金属NPよりも優れています[8、9、10、11、12]。 AuNPの細胞毒性および遺伝子毒性効果は、その形状、サイズ、電荷、濃度、相互作用時間などに関連しています[12、13、14]。したがって、臨床現場でナノマテリアルを使用するには、それらの潜在的な毒性と人間の健康への影響に関する情報が必要です。
現在、標的療法で大きな成功を収めているにもかかわらず、標的組織でのAuNPの選択的送達の問題は未解決のままです。いくつかの研究は、異なる起源の上皮細胞によるNPの異なる取り込み率を指摘しています[15、16]。しかし、AuNPの組織選択的ターゲティングを達成するのに役立つ可能性があるとしても、この現象を説明するための調査は不足しています。異なる上皮間の解剖学的または生理学的差異は、AuNPの取り込みおよび輸送速度の差異を説明する可能性があります。特に、取り込み速度は、細胞の原形質膜特性、ナノ粒子の細胞表面糖タンパク質およびプロテオグリカンへの結合、ならびに細胞の小胞輸送能力によって影響を受ける可能性があります[17]。したがって、ナノ粒子と標的細胞との排他的な選択的相互作用の不可能性を考慮に入れて、癌治療の望ましくない結果を回避するための正常細胞および発癌性細胞との相互作用の特徴の比較研究が話題になっています[8,9,10]。
インビボモデルはナノ粒子の生物学的毒性を評価するために価値がありますが、細胞培養モデルは前臨床の生理学的および毒物学的研究に非常に役立ちます。現在、細胞培養は生物学、医学、獣医学、バイオテクノロジーのさまざまな分野で広く使用されています。細胞培養を使用することで、生物のレベルで研究することが困難で、時には不可能な生物学的プロセスを探求することができます。細胞培養の重要な役割は、多くのワクチン、試験システム、および生物活性物質の生産におけるバイオテクノロジーで果たしています。細胞培養は、さまざまな病因の疾患を診断するために使用され、新しい薬理学的、治療的、美容的薬剤、および食品添加物をテストする際のテストオブジェクトとして使用されます[18]。
細胞培養モデルに関するこの研究では、連続的および発癌性の細胞株起源の上皮細胞のAuNPの効果の特徴を調べることを試みました。上皮細胞の単層培養SPEV(胚性ブタ腎臓上皮接種株)およびHT-29(結腸癌細胞株)細胞は、AuNPsによる抗腫瘍療法が適用される場合、正常および癌上皮組織のモデルと見なすことができます。 AuNPの細胞毒性を検証するために、接着、増殖、壊死/アポトーシス、多細胞スフェロイドなど、いくつかの従来の細胞毒性アッセイが採用されました。
メソッド
SPEV細胞の培養
SPEV細胞は、5%FCS( v )を含むDMEM(Sigma、USA)のプラスチックフラスコで培養しました。 / v )(HyClone、USA)ペニシリン/ストレプトマイシン(PAA、オーストリア)およびアンホテリシンB(5μg/ ml)(5%CO 2 、湿度95%)[19]で報告されています。播種濃度は0.5–2×10 4 でした セル/ cm 2 。培地は2日ごとに交換しました。細胞は100%コンフルエンスで継代されました[20]。 SPEV細胞株は、培養中の細胞変性の証拠なしに、連続継代中に単層の初期形態学的構造を成長させ、保存しました。
HT29細胞の培養
HT29細胞は、10%FCS( v )を含むRPMI-1640培地(Sigma、USA)のプラスチックフラスコ(Nunc、デンマーク)で培養されました。 / v )(HyClone、USA)2 mM L-グルタミン(Sigma、USA)および40 mg / mlゲンタマイシン(Sigma、USA)を標準状態(5%CO 2 )で添加 、湿度95%)[21]。最適な細胞密度は0.5〜4.0×10 4 でした セル/ cm 2 。細胞は、ウクライナのRE Kavetsky Institute of Experimental Pathology、Oncology and RadiobiologyNASのヒトおよび動物組織からの細胞株バンクから提供されました。
AuNPを使用した操作
AuNPは、ロシア科学アカデミーの植物および微生物の生化学および生理学研究所から提供されました。 AuNPはクエン酸塩法[22]によって合成されました。ナノ粒子の平均サイズは15nmでした。金の初期濃度は57μg/ mlでした。暗視野電子顕微鏡の結果、15 nm AuNPの画像、および15 nm AuNPの吸光スペクトル(b)を図1に示します(図1a;サイズ分布の注-図)[23]。 AuNPは、37°Cでの受動拡散によって細胞に導入されました。調査した濃度は1、3、6、および12μg/ mlでした。同じ条件下でAuNPを含まないセルをコントロールセルとして使用しました。
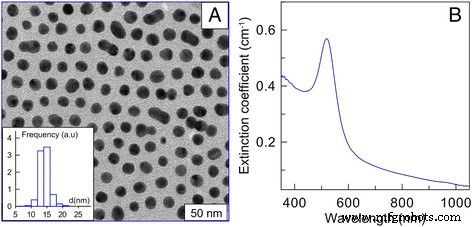
暗視野電子顕微鏡の結果、 a 15 nm AuNPの画像(注-サイズ分布の図)、 b 15 nmAuNPの減光スペクトル
接着および増殖細胞
細胞培養の形態機能状態は、接着特性と増殖活性によって判断されました。 SPEVおよびHT29細胞の接着特性は、倒立顕微鏡を使用して視覚的に評価されました。付着および平坦化した細胞の数は、培養開始後30、60、120、180、および1440分でカウントされました。
SPEVおよびHT29細胞の増殖動態を、1〜4日間研究しました。研究された培養物中の細胞数の増加を調査条件まで調べるために、それらをプラスチックから酵素的に(1:1(0.25%トリプシン溶液:EDTA、PAA))剥離し、細胞数を数えた。培養細胞の総数は、ゴリヤエフのチャンバーで従来の方法でカウントされました。
アポトーシス/壊死プロセス
AuNPに曝露されたSPEVおよびHT29細胞のアポトーシスおよび壊死プロセスは、FACS Calibur Becton-Dickinsonを使用して、アネキシンV(BD、米国)、7-アミノアクチノマイシン(7AAD)(BD)色素を使用して4日間で調査されました。結果はWinMDIv.2.8ソフトウェアで分析されました。
多細胞スフェロイド
多細胞スフェロイド(MS)は、調査対象の細胞の移動と凝集の可能性に対するAuNPのinvitroでの影響を推定するために生成されました。 SPEVおよびHT29細胞のスフェロイド(3-D)モデルシステムは、[24]によって報告され、私たちの研究室[25]で修正された従来の方法で培養されました。簡単に説明すると、トリパンブルーと同数の細胞(5×10 4 )を使用して細胞懸濁液をカウントしました。 セル/ cm 2 )完全に補充された培地に植えられた。 MSの生成は、1%寒天でコーティングされた24ウェルプレートで0.24%のカルボキシメチルセルロース(CMC)を使用して、回転(80 rpm)で24時間細胞培養を行うことで達成されました。その後、3次元細胞培養は標準的な条件で維持されました。 MSのサイズと数のAuNP濃度への依存性を調査するために、AuNPの存在下でMSを生成しました。プレートを一定に回転させながら、さらに48時間培養しました。次の段階では、Carl Zeiss Stemi2000顕微鏡を使用した暗視野法によってマイクロ写真画像を作成しました。 MSの形態は、Axio Vision Release 4.7プログラム(Zeiss)の助けを借りて研究されました。このプログラムは、細胞凝集体の幾何学的寸法を測定することを可能にします。次に、ファイルに含まれていたすべてのMSのボリュームを計算しました。次の式で使用されました: V =0.4× a × b 2 、ここで a および b -MSの幾何学的直径[24]。統計分析のために、すべての細胞凝集体を1×10 -4 のサイズでグループ化しました。 mm 3 〜1×10 −2 mm 3 1×10 -3 の増分で mm 3 。各グループのMS数とMSボリュームの中央値を推定しました。
統計分析
分散分析と学生の t の単一因子分析 テストは、ソフトウェアパッケージStatistica8を使用したデータの統計処理に使用されました。有意水準は0.05でした。結果は、平均値と標準誤差(M±SE)として表示されます。
結果
SPEVおよびHT-29細胞の接着に対するAuNPの影響
細胞接着は細胞の機能状態の指標であり、培養のさらなる成長に必要です。接着が終了すると、細胞は平らになり、適切な形態になりました。 SPEV細胞の接着特性を図2に示します。
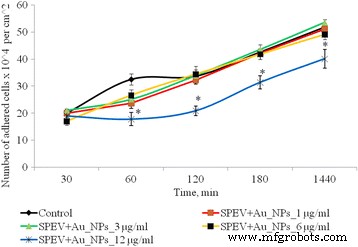
AuNPの曝露後のSPEV細胞の接着のダイナミクス、* p ≤0.05は、コントロールと比較して有意です
1、3、および6μg/ mlのAuNPを使用してSPEV細胞を1時間培養した後、付着した細胞の数は対照値よりも少なくなりました。これらの濃度のAuNPを含むサンプルの平坦化された細胞の割合は、コントロールと有意差はありませんでした。 12μg/ mlのAuNPと1時間インキュベートした後、接着が遅くなりました。平方センチメートルあたりの付着細胞の数は、コントロールと比較して1.8倍減少しました。この付着傾向は、すべての試験期間にわたって持続した。 24時間の観察後、付着した細胞の数は、コントロールと比較して1.3倍少なくなりました。同時に、低濃度(1および3μg/ mlと腫瘍細胞(HT29))のAuNPのインキュベーションは、接着細胞の量に有意な影響を与えませんでした。AuNP濃度を6および12μg/ mlに増やすと、接着画分中の腫瘍細胞数はそれぞれ1.16倍と1.28倍(図3)。得られたデータはいくつかのプロセスの影響を受ける可能性があります。1つは腫瘍との両方の接着画分に対するAuNPの細胞静止/細胞毒性効果です。胚性細胞株は、細胞死、アポトーシスへの移行、または壊死を引き起こします。もう1つのプロセスは、AuNPの影響下での細胞接着の低下と、細胞の浮遊画分への移行です。特に、両方のプロセスを同時に実現できます。そして、それぞれが接着画分の生細胞数の減少に寄与する可能性があります。
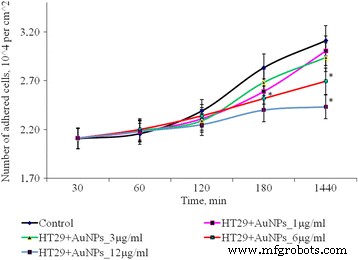
AuNPs、* p の曝露後のHT29細胞の接着のダイナミクス ≤0.05は、コントロールと比較して有意です
SPEVおよびHT-29細胞の増殖に対するAuNPの影響
1〜12μg / mlの濃度範囲内のAuNPが、SPEV細胞培養の増殖過程に及ぼす影響を調べました(図4)。 1、3、および6μg/ mlのAuNPで培養した2〜4日目では、細胞数はコントロールと有意な差はありませんでした。 3および6μg/ mlのAuNPで培養した4日目に、この指数は対照と比較してそれぞれ1.15倍および1.23倍減少しました。 SPEV培養では、対照と比較して、12μg/ mlのAuNPを用いた培養の1.5倍(2日目と3日目)および4日目で1.15倍の細胞数の減少が観察されました。したがって、AuNP濃度12μg/ mlは、観察された期間内に細胞増殖を遅らせました。
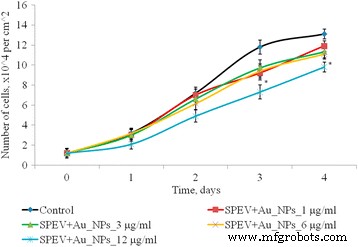
AuNPの曝露後のSPEV細胞の増殖、* p ≤0.05は、コントロールと比較して有意です
単層培養におけるHT29細胞の数に対する1〜12μg / mlの濃度のAuNPの影響を図5に示します。インキュベーションの最初の3日間、コントロールおよび存在下の細胞数AuNPの数は統計的に異ならなかった。培養4日目に、2D培養における細胞数の用量依存的な減少が認められました。したがって、4日間の培養後、低濃度のAuNP(1および3μg/ ml)の場合、HT29細胞の数はコントロールと比較して有意差はありません。しかし、より高いAuNP濃度(6〜12μg / ml)では、HT29細胞数は対照よりもそれぞれ1.33倍と1.44倍低かった。
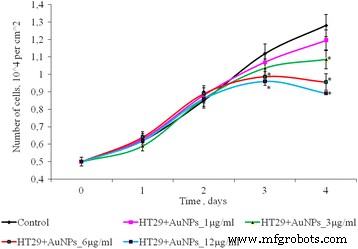
AuNPの曝露後のHT29細胞の増殖、* p ≤0.05は、コントロールと比較して有意です
SPEVおよびHT-29細胞のアポトーシス/壊死プロセスに対するAuNPの影響
AuNPの存在下でSPEVおよびHT-29細胞を、標準的な条件下で4日間培養しました。 1および3μg/ mlのAuNPを使用したSPEVおよびHT29細胞の培養、およびアポトーシス/壊死プロセスの指標は、コントロールと有意な差はありませんでした(表1および2)。
<図> <図>6〜12μg / mlのAuNPで培養すると、アネキシンV + / 7AAD +、アネキシンV- / 7AAD +、およびアネキシンV + / 7AAD細胞の割合が増加し、生細胞の割合が減少しました。アネキシンV + の数 / 7AAD + SPEVセルは、コントロール値より7.8±0.7%高かった( p ≤0.05)12μg/ mlのAuNPを使用。アネキシンV + の数 / 7AAD + HT 29セルは、コントロール値より3.2±0.4%高かった( p ≤0.05)6μg/ mlのAuNPを使用し、4.8±0.6%( p ≤0.05)12μg/ mlのAuNPを使用。
SPEVおよびHT29細胞からの多細胞スフェロイドの生成に対するAuNPの影響
多細胞スフェロイド(MS)のサイズと数のAuNP濃度への依存性を判断するために、48時間の間にさまざまな濃度のAuNPでMSを生成しました。私たちのデータは、HT29およびSPEV細胞が、微小環境の同じ条件下で多細胞スフェロイドを形成するさまざまな能力を示しました(図6および7)。
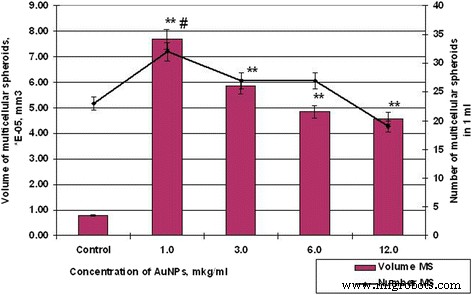
AuNPとのインキュベーション後のMS細胞SPEVの数と量、# p ≤0.01(MS数の場合)。 ** p ≤0.01(MSのボリュームの場合)
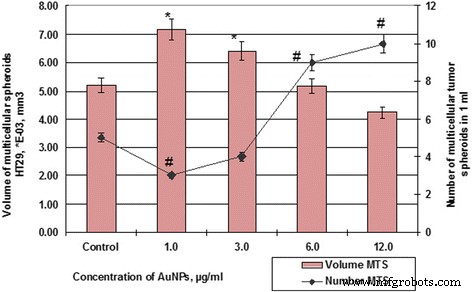
AuNPとのインキュベーション後のMS細胞HT29の数と量。 # p ≤0.01(MS数の場合)。 ** p ≤0.01(MSのボリュームの場合)
したがって、HT29細胞の48時間の対照サンプルが、平均体積5.19×10 -3 のスフェロイドを形成した場合 mm 3 、SPEVセルの平均体積スフェロイドは0.79×10 -5 でした mm 3 。同時に、HT29およびSPEVセルに対するAuNPの影響は同じ傾向でした。細胞微小環境におけるAuNPの存在は、両方の培養物における多細胞スフェロイドの形成を刺激しました。したがって、AuNPの濃度が1および3μg/ mlの場合、SPEVのMSの量はコントロールと比較してそれぞれ9.7倍および7.4倍増加し(図6)、同じAuNP濃度もMSの量の増加を刺激しました。 HT29はそれぞれ1.4倍と1.2倍です(図7)。
AuNP濃度をさらに上げると、両方の培養でMSの平均量が減少しました。 AuNP濃度が1から12μg/ mlに上昇すると、HT29MSの量が7.18×10 -3 に減少しました。 mm 3 〜4.24×10 -3 、コントロールによると、1.69回で。 SPEVの場合、AuNPの濃度を1から12μg/ mlに増やすと、MSの量は7.69から4.58×10 -5 に減少しました。 mm 3 、コントロールによると、1.68回で。ただし、AuNP濃度の増加は、MSの量の減少と一致し、HT29細胞の培養におけるスフェロイドの数の増加と相関していました(図6および7)。 HT29 MSの数は、AuNP濃度が1から12μg/ mlの場合、視野あたり3から10に増加しました。同時に、SPEVMSの数はそれぞれ32から19に減少しました。
得られたデータ(図6および7)は、AuNPが細胞間システムの凝集相互作用に影響を与えることができることを示しています。私たちのデータは、低濃度のAuNP(1〜3μg / ml)が胚細胞と腫瘍細胞の両方の多細胞スフェロイドの形成を刺激したことを示しています。ただし、AuNP濃度が高い(6〜12μg / ml)と、懸濁液中の細胞に細胞毒性と抗凝集効果の両方がありました。このプロセスは、平均量が減少した多数のHT29MSの形成に貢献しました。 SPEVに関しては、高濃度のAuNPは細胞増殖抑制効果を持ち、接着画分の細胞数と懸濁液中のMSの数を減少させる可能性があります。以前、著者らは、カーボンナノ粒子が細胞の基質への接着を減少させ、懸濁液への細胞の移動を刺激し、多細胞スフェロイドの形成をもたらすことを報告しました[25、26]。文献には、アクチン/ミオシンマイクロフィラメントの構造を破壊し、細胞の増殖、接着、および分化を減少させるAuNPの能力に関するデータがあります[27]。私たちのデータはこの仮定を確認しました。
ディスカッション
上皮細胞の連続的および発癌性細胞株起源の増殖、壊死/アポトーシス、および多細胞スフェロイドの形成に対するAuNPの効果を評価した。 6〜12μg / mlのAuNPは、SPEVおよびHT29細胞の数を減らし、アポトーシスおよび壊死の初期および後期の細胞数を増加させることが示されました。低濃度のAuNPは、HT29およびSPEV細胞による多細胞スフェロイドの形成を刺激します。ただし、AuNP濃度が高いと、懸濁液中の細胞に対して細胞毒性と抗凝集効果の両方がありました。 AuNPの作用に対する大きな感度は、SPEV細胞(12μg/ ml)と比較してHT29(6μg/ ml)の系統によって示されました。
細胞形態と細胞骨格に対するAuNPの影響は最近注目を集めたばかりであり、根本的なメカニズムと今後の結果は詳細に調査されていません[28、29、30]。この点で、すべての新規AuNPタイプが、時間の関数としてエンドサイトーシスの取り込み経路と細胞内局在を評価することが重要です。さまざまなタイプのAuNPの場合、影響は細胞内AuNP濃度に依存し、一過性であると説明されています。細胞分裂が繰り返されると、細胞内AuNP濃度は指数関数的に減少し、影響は観察されなくなります。また、AuNPのエンドソーム脱出の可能性を評価する必要があります。細胞骨格の欠陥はAuNP濃度に明らかに依存していると説明されているため、影響を与えることなく最大の細胞負荷容量を評価するために、広範囲の粒子をテストする必要があります。さらに、細胞骨格は多くの細胞内シグナル伝達経路にも関与しているため、AuNPが細胞骨格の破壊を誘発して二次的影響をもたらすかどうかはまだ調査されていません[31]。
NPは特定の物理的寸法を持っているため、NPが占める細胞内容積は、細胞形態の変化を引き起こしたり、細胞骨格ネットワークの構造に影響を及ぼしたりする可能性があります[28、29、31]。後者の影響はまた、細胞のエンドサイトーシス様式に対するNPポーズの高い要求に起因する可能性があります。 AuNPは、A549ヒト癌性肺細胞などのいくつかの細胞型の形態に大きな影響を与えることが報告されています[32]。 AuNPは、ヒト皮膚線維芽細胞のアクチン原線維に濃度依存性の影響を与えることも報告されています[33、34]。 Mironava etal。 [35、36]はさらに、アクチンまたはβ-チューブリンタンパク質の発現レベルは影響を受けなかったものの、AuNP曝露時間、濃度、およびNPのサイズの関数として細胞骨格フィラメントが破壊されることを示しました。
使用される細胞の種類も非常に重要です。なぜなら、密接に関連している場合でも、同じ種類のナノ材料に対しては、異なる細胞の種類がまったく異なる反応を示す可能性があるからです[37、38]。好ましくは、NPの(将来の)生物医学的応用に最も関与する細胞型(例えば、上皮細胞、内皮細胞)、または異なる胚葉に由来する複数の細胞を試験する必要があります。細胞毒性の影響を調査する場合、異常な結果につながる可能性があるため、がん細胞タイプの使用は最小限に抑える必要があります[39]。癌細胞にはいくつかの特定の特徴があり、細胞内シグナル伝達経路が変化しており、増殖をアップレギュレートし、細胞の生存能力を維持するため、NPを介した細胞毒性効果が起こりにくくなります。
私たちの意見では、細胞の表面官能基(膜貫通タンパク質など)へのAuNPの結合は、可逆的または不可逆的であり、一時的または永続的な構造的損傷をもたらす可能性があります[40、41]。細胞の生体力学的特性(硬度や弾性など)、接着性、および表面の電気的特性の変化の潜在的な影響が認識されます。したがって、硬度または弾性の変化は、表面構造の柔軟性、細胞分裂のための機械的エネルギーの生成、および細胞の運動性に影響を与える可能性があります。接着性に関しては、細胞の微小環境は通常、細胞が周囲に接着することを可能にする特定の分子を含む細胞外マトリックスで構成されています[42]。表面電荷は、間違いなく細胞とその周囲の間の相互作用において重要な役割を果たします。
他の著者も、NPがミトコンドリアに優先的に局在し、酸化ストレスを引き起こし、構造的損傷を増強することを報告しました[40]。パンらによる最近の記事。 1.4 nmのAuNPは、Hela細胞の酸化ストレスとミトコンドリアの損傷を介して壊死を誘発すると説明しています[43]。生分解時に細胞培地にナノ粒子が蓄積すると、細胞小器官が破壊されたり、遺伝子変異が引き起こされたりする可能性があるため、安全ではありません。
アポトーシス中に細胞で発生する変化は、ほとんどの細胞タイプで類似しています。アポトーシス細胞では、原形質膜の脂質組成に変化があります。ホスファチジルセリンが二重層の細胞質部分から外側に移動し、カスパーゼカスケードの活性化、クロマチン凝縮、ミトコンドリアの電子伝達系の障害を引き起こし、最終的にATP合成を停止させます。プログラム細胞死は、遺伝的障害、化学的または物理的要因への曝露、および細胞の他の変化から生じる受容体を介した生理学的刺激によって引き起こされる可能性があります。この効果は、6〜12μg / mlのAuNPで観察されました。
多細胞凝集体(スフェロイド、胚様体)は、単層増殖細胞と組織培養の間の断続的なレベルを表します。スフェロイドは、細胞の3次元成長と組織化、細胞間の相互作用、および微小環境条件、たとえばAuNP濃度、増殖の強度、細胞の接着性および微小凝集体の形成の影響の客観的モデルです。 MS形成は、胚細胞株と同様に腫瘍に関しても確立された培養方法です[24、44、45]。私たちの研究では、スフェロイドの形成と成長は、人工細胞外マトリックスの一部としてCMCを追加し、表面への細胞接着を抑制し、細胞凝集を刺激する1%寒天による表面コーティングによって達成されます。これらの条件では、限られた細胞塊の成長または胚性細胞の自発的分化により、凝集体が中枢壊死を形成するまでMS培養を実現できます。
文献には、AuNPと結腸癌細胞株および胚細胞株との相互作用に関する情報があります[46、47]。これらのデータによると、非常に低濃度のAuNPへの曝露でさえ、細胞増殖、分化、およびアポトーシス細胞死にストレスを与えることにより、ヒト胚性神経前駆細胞およびHT29に有害な影響を与える可能性があります。
AuNPの効果は、G0 / G1期の蓄積、S期およびG2 / M期の枯渇、ならびにヒト口腔扁平上皮癌細胞(HSC-3)のATPレベルの低下に基づくというデータが公開されています[48]。細胞周期の調節は、2D培養での基質と細胞の浮遊画分への移動と細胞の焦点接触の違反、および3D培養でのギャップ結合での細胞間接触の阻害によって対処される可能性があります[48、49、50]。ナノサイズのAuNP(15 nmに近い)のため、細胞の凝集の中心にはなりません。同時に、AuNPの細胞膜へのインターカレーション[51]、細胞膜のAuNPゼータ電位への影響[32]、および細胞間/細胞間接触の形成への影響は、明らかに壊死/アポトーシスのメカニズムを引き起こす可能性があります、細胞毒性効果、および細胞周期停止。細胞と基質との焦点接触の違反および浮遊画分への細胞移動は、細胞周期調節の方法である[48、49]。低濃度のAuNPは、細胞に対して統計的に有意な細胞毒性効果を発揮しませんでした。ただし、AuNP濃度が高いと、懸濁液中の細胞に対して細胞毒性と抗凝集効果の両方がありました。このプロセスは、平均量が減少した多数のMSの形成に貢献しました。 AuNPは細胞の凝集接触にくさびを入れ、それらを危険にさらすと考えられます。したがって、SPEVおよびHT29細胞株に対するAuNPの効果に関する実験は、AuNPを介した癌治療の開発におけるそれらのさらなる応用をサポートします。
しかし、invivo動物実験に対する抗がん効果を確認するために将来の研究が必要になるでしょう。それにもかかわらず、私たちの深い確信は、物質の性質とその可能性のある負の影響を知っていれば、AuNPの有害な影響を回避し、それらの正のバイオテクノロジーの可能性を利用できるということです。私たちの調査は、抗がん治療の文脈で効果的な材料に非常に確実に適用でき、医学にとって最大の利点があります。
結論
私たちの結果は、AuNPがSPEVおよびHT29細胞で用量依存的な細胞毒性を誘発するという概念を支持しています。さらに、このレポートは、6〜12μg / mlの濃度の15nm AuNPが、SPEVおよびHT29細胞の増殖を減少させ、アポトーシスおよび壊死の初期および後期段階で細胞数を増加させたことを初めて示しています。また、低濃度のAuNP(1〜3μg / ml)が多細胞スフェロイドの形成を刺激することが示されました。ただし、AuNP濃度が高いと、懸濁液中の細胞に対して細胞毒性と抗凝集効果の両方がありました。 AuNPの作用に対する大きな感度は、SPEV細胞(12μg/ ml)と比較してHT29(6μg/ ml)の系統によって示されました。
ナノマテリアル
- 化学療法センサー用の金ナノ粒子
- 改善された診断および治療用途のための多機能金ナノ粒子:レビュー
- 癌治療のためのナノ粒子:現在の進歩と課題
- コバルトをドープしたFeMn2O4スピネルナノ粒子の調製と磁気特性
- ICAをロードしたmPEG-ICAナノ粒子の調製とLPS誘発性H9c2細胞損傷の治療におけるそれらの応用
- invitroおよびinvivoでのグラフェンおよび酸化グラフェンのバイオセーフティおよび抗菌能力
- 光熱治療および光音響イメージングのためのポリピロール被覆鉄-白金ナノ粒子の合成およびinvitro性能
- 6-メルカプトプリンとニューロン透過性ペプチドで修飾された金ナノ粒子によるSH-SY5Y細胞増殖の促進
- ナノ粒子の跳ね返りに及ぼす弾性剛性と表面接着の影響
- ポリマーナノコンポジットのヤング率に対するナノ粒子の凝集/凝集の影響を研究するための2段階の方法論
- 強化された細胞内在化のためのFe3O4磁性ナノ粒子の葉酸とgH625ペプチドベースの機能化の比較