


小さな希土類フッ化物ナノ粒子は、電気極性相互作用を介して腫瘍細胞の成長を活性化します
要約
ナノ粒子と膜貫通シグナル受容体の間の局所的な細胞外相互作用は、癌細胞の成長を十分に活性化する可能性があります。ここでは、小さなLaF 3 およびPrF 3 DMEM + FBS懸濁液中のナノ粒子は、3つの異なるヒト細胞株(A549、SW837、およびMCF7)で腫瘍細胞の増殖を刺激しました。ナノ粒子のサイズ分布、AKTおよびERKシグナル伝達経路の活性化、および生存率テストは、小さなサイズのナノ粒子(<10 nm)の集合体の相乗作用を介したインテグリンおよびEGFRのリガンド接着結合部位の機械的刺激を示しました。小さなサイズのナノ粒子はEGFRの活性化とよく関連している可能性がありますが、インテグリンとナノ粒子の相互作用は多面的な問題のままです。理論的なモチーフは、必要なpN力のスケール内で、各リガンド接着結合部位が電気双極子相互作用を介して小さなサイズの誘電体ナノ粒子によって活性化できることを示しています。活性ナノ粒子のサイズは、リガンド接着結合部位とナノ粒子の表面電荷の量、およびそれらの間の分離距離によって指定されたままでした。電気双極子力の極成分は、ナノ粒子のサイズの2乗に反比例したままであり、小さなサイズの誘電体ナノ粒子のみが電気双極子相互作用を介して癌細胞の成長を刺激する可能性があることを示しています。この研究は、癌細胞のさまざまな細胞骨格ストレスモードの認識に貢献しています。
背景
腫瘍形成は、ゲノムの変化を伴う多次元の問題です。また、足場と細胞骨格構造の間の細胞外マトリックス(ECM)相互作用によっても活性化され[1,2,3,4]、インテグリンと同様に、ゲノムプログラミングを変更できるマルチパート細胞力からのメカノセンサーのストレスによって発現されます[5]。腫瘍微小環境とECM足場との相互作用は、通常、細胞の膜接着斑タンパク質と膜貫通シグナル受容体(TSR)、上皮成長因子受容体(EGFR)、血管内皮増殖因子(VEGFR)、または神経成長因子受容体(NGFR)を活性化します。メカノセンサーは、リン酸化反応の雪崩を引き起こすことにより、細胞の細胞外活性ドメイン[6,7,8,9]と細胞内F-アクチンフィラメント間のシグナルトランザクションを介して腫瘍細胞の成長を調節します。
タンパク質のコンフォメーション変化とTSR経路の励起には、pN力の範囲内、そして確かにnNゲージより下にある活性化力が必要です[10]。ランダムな機械的ストレスとアクティブな化学親和力の強さに加えて、ナノ粒子(NP)と細胞膜のタンパク質間の結合効率(結合の強さ)は、短距離または長距離の電気極性または他のタイプの分散相互作用を介して調整できます。 NPの限られた表面積では、生物学的に活性であるのに十分長い間、特定の数のタンパク質のみを付着させることができ[11]、生物学的環境との空間に限定された局所的相互作用が一連の多様な細胞機能の原因であると認識されましたルート[12]。その結果、タンパク質とNPの相互作用からのシグナルトランザクション経路は、NPの安全性の問題をシグナルアウトしています[11、13]。
NPからの細胞における好ましいまたは不利な反応はタイプ特異的であるため[11]、NPと生物学的マークの間のリンクはケースバイケースで確立されるべきです[14、15]。
NPに曝露された腫瘍細胞からの矛盾する結果は、切除または腫瘍増殖効率またはNPの可変毒性レベルのいずれかについても[16、17]、安全性の問題を浮き彫りにしている。それにもかかわらず、進歩にもかかわらず、今日、NPが真核細胞と相互作用する特定の経路に関する知識が不足しており、普遍的なNP治療アプローチの特定が不可能になっています。異なるサイズのNPと多様な表面化学は通常、NP-膜受容体結合やTSR活性などの細胞応答をそらすため、NPの毒性は、界面活性剤の形態、帯電状態、ECM中のタンパク質とナノ材料の濃度と組成に関連しています。 [18,19,20,21]そして最後にNPと細胞表現型の間の分子間結合の強さ[22]。
シリカ、金のNP、およびカーボンナノチューブに曝露された黒色腫および頸部癌の以前の研究では、NPのサイズが腫瘍細胞の成長を選択的に活性化することが認識されていました[23、24、25、26、27]。ヒトSK-BR-3癌細胞株のTSRシグナルと、修飾された金および銀のNPのサイズとの相関関係は、2〜100 nmのサイズのNPがシグナル伝達を再形成したものの、細胞が40と相互作用するとアポトーシス活性に大きな違いが得られることを示しました。 –50nmサイズのNP [26]。最近、金NPのサイズを5nmから40nmに変更すると、A549および95D癌細胞株の増殖速度が調整されたと考えられます。具体的には、5 nmサイズのNPは両方の細胞タイプの増殖を抑制しましたが、約10nmサイズのNPは細胞増殖に影響を与えませんでした[27]。同様に、SiO 2 に曝露されたA549およびTHP-1細胞 NPはサイズ依存性の細胞毒性を示し、15nmサイズのNPも高い細胞毒性レベルと相関していた。それどころか、60nmサイズのNPはより低い毒性を示しました。最後に、200 nmサイズのNPは、ERK1 / 2の活性化を通じて幹細胞の成長を増加させましたが、2〜4μmサイズのNPは、さまざまなシグナル伝達経路を活性化することができました[28]。小型のNPは、EGFRを結合し、プロテインキナーゼB(AKT)と細胞外シグナル調節キナーゼ(ERK)のシグナルトランザクション経路をオンにして、細胞の成長を促進します。
希土類ナノ粒子(RE-NP)は、金属イオン依存性接着部位(MIDAS)、MIDASへの調整(ADMIDAS)、相乗的金属イオン結合部位(SyMBS)、リガンド接着結合部位(LABS)などの特定のドメインとも相互作用する可能性があります。 、ανにあります β 3 サブユニットまたは他のインテグリンサブユニット[29、30]。
同様に、RE-NPは、腫瘍NPの相互作用にさらなる柔軟性をもたらします[31、32、33、34]。セリアNP(ナノセリア)は、さまざまなラジカルによる細胞損傷に対する保護作用を示しましたが[35]、低濃度レベルの修飾セリアNPは、AKT / ERKシグナル伝達経路の活性化を介してアポトーシスを減少させることにより肝細胞癌細胞増殖を大幅に促進しました[36]。通常、細胞を取り巻くNPの集合は、細胞骨格ストレスの影響を受けやすく、NPとメカノセンサー間の化学的、ナノ熱力学的(Hill)[37]、エントロピーまたは電気双極子相互作用に対しても同様に立ちます。ただし、これまで、NP、TSR、およびセル間の相互作用を理解するためのケーシングはあいまいであり、利用できません。
原則として、RE化合物の強いイオン特性は、電気的相互作用を介して細胞のメカノセンサーを刺激するはずです。また、REイオンはさまざまなアプリケーションで広く使用されているため、適切な公衆衛生保護プロトコルを示唆するために、腫瘍細胞の成長への潜在的な寄与を調べることが重要です。フッ化ランタン(LaF 3 )およびフッ化プラセオジム(PrF 3 )は、蛍光灯、放射色ガラス、光ファイバー、エナメルのアプリケーション、および電極で使用されます。 LaF 3 特定の種類のガラス、リン光ランプコーティング、水処理、および触媒で精巧に作られています。また、市販のフッ化物ガラス(ZBLAN)の必須成分であり、フッ化ユーロピウムと混合して、光通信や、赤外線透過率の高いイオン選択性フッ化物電極の結晶膜として使用されます。同様に、PrF 3 映画産業向けのカーボンアークランプ、スタジオ照明、プロジェクターライトにも採用されています。プラセオジムをドープしたフッ化物ガラスは、シングルモードファイバ光増幅器にも使用されています。
これにより、この研究は、小さなサイズのRE-NPが電気双極子相互作用を介して腫瘍細胞の成長を刺激する能力を持っていたことを示しています。
この記事は3つのセクションで構成されています。まず、動的光散乱(DLS)、原子間力顕微鏡(AFM)、透過型電子顕微鏡(TEM)、X線回折(XRD)、2次元高速を適用して、NPのサイズ分布、相互作用、および形状を分析します。フーリエ変換(2D-FFT)分析および真空紫外線分光法(VUV 110–180 nm)。次に、3つの異なるヒト癌細胞株(A549、SW837、およびMCF7)の増殖レベルと、サイズ分布およびLaF 3 の濃度との相関関係 およびPrF 3 NPが確立されます。最後に、メカノセンサーを活性化し、続いて腫瘍細胞を増殖させるために必要な1 pNの力の制限内で、腫瘍細胞の生存率は、1つのRE-NPと1つのLABS間の電気双極子相互作用の理論的モチーフに適合します。この研究は、さまざまな種類の細胞骨格ストレスの特定と分類、およびNPと癌細胞の機械感覚との相互作用に貢献しています。
結果
NPのサイズと構造
まず、DLS、AFM、TEM、XRD、FFT、VUV分光法、および t 液体懸濁液中のRE-NPのサイズ分布を抽出するために検定統計量が適用されました(図1、2、3、4、5、6、および7)。次に、細胞の生存率試験とウエスタンブロッティング(Wb)アッセイを使用して、RE-NPによる特定のメカノセンサーの活性化を特定しました。
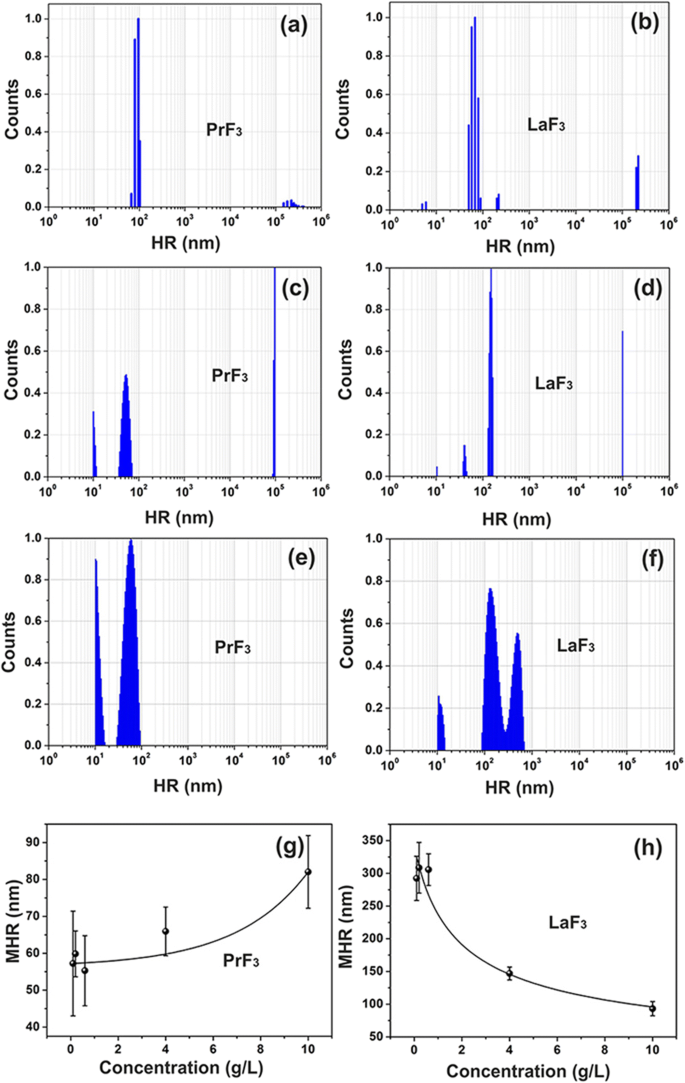
RE懸濁液のDLSサイズ分布スペクトル。 a、b PrF 3 およびLaF 3 水中のNP(5g / L)。 c、d PrF 3 およびLaF 3 DMEM + FBS中のNP(5g / L)。 e、f PrF 3 およびLaF 3 DMEM + FBS中のNP(0.1g / L)。 g、h PrF 3 の標準偏差を持つ平均流体力学的半径(MHR) およびLaF 3 異なる濃度レベルでのDMEM + FBSのNP
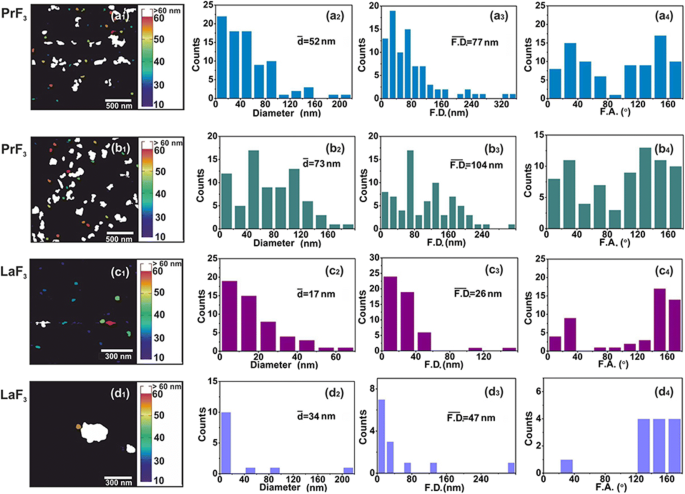
AFMデジタル( x 、 y )RE-NPのサイズヒストグラム。 ( a1–d1 )PrF 3 (2×2μm 2 )およびLaF 3 (1×1μm 2 )DMEM + FBS懸濁液中のNP。 ( a2–d2 )( x 、 y )RE-NPの平均等円面積直径\(\ left(\ overline {d} \ right)\)のサイズヒストグラム。 (a3–d3)( x 、 y )RE-NPのフェレット領域の直径\(\ left(\ overline {\ F.D.} \ right)\)のサイズヒストグラム。 ( a4–d4 ) x を基準にしたフェレット角度(F.A.)ヒストグラム -軸。 RE-NPは、±(44–60°)
の間の2つの主要な方向に沿って方向付けられました。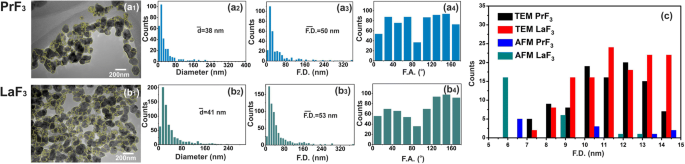
TEMデジタル( x 、 y )RE-NPのサイズヒストグラム。 ( a1、b1 )RE-NPのTEM画像。黄色の線は、RE-NP間の2D境界を示しています。 ( a2、b2 )( x 、 y )RE-NPの平均等円面積直径\(\ left(\ overline {d} \ right)\)のサイズヒストグラム。 ( a3、b3 )( x 、 y )RE-NPのフェレット面積直径\(\ left(\ overline {\ F.D.} \ right)\)のサイズヒストグラム。 ( a4、b4 ) x を基準にしたフェレット角度(F.A.)ヒストグラム -優先方向が±(44–60°)の軸。 ( c )4μm 2 のAFM画像とTEM画像の両方から抽出された小型および小型のRE-NPのフェレット直径のサイズヒストグラム エリア
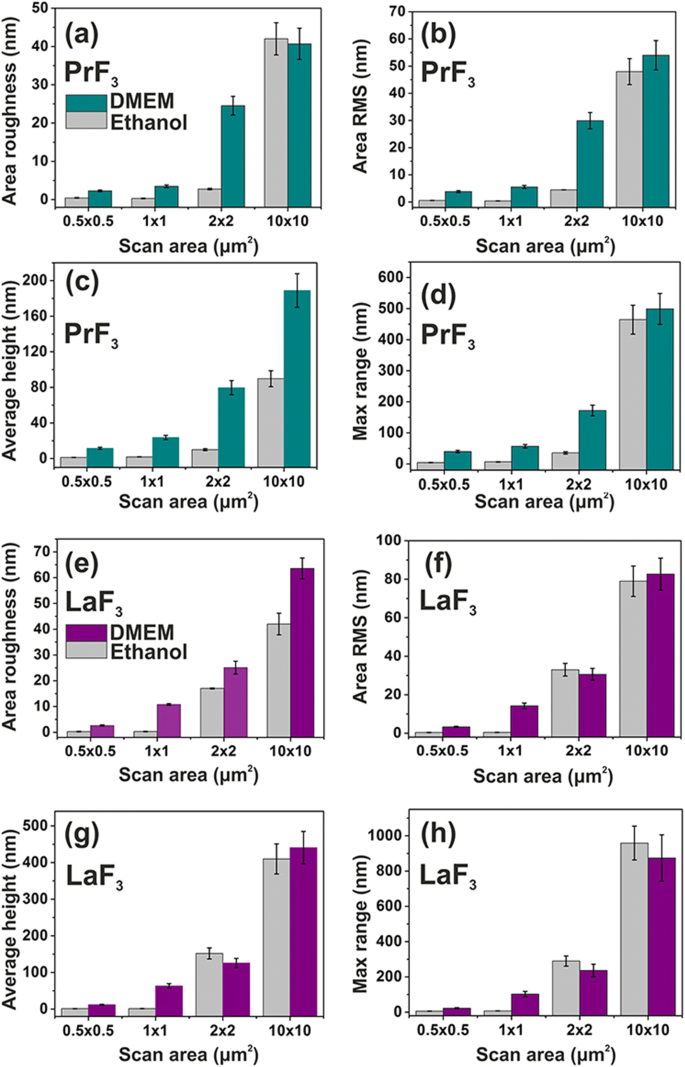
PrF 3 の表面パラメータヒストグラム(z-height) およびLaF 3 さまざまなスキャン領域のDMEM + FBSおよびエタノール中のガラス基板上のRE-NP。 a 、 e エリアの粗さ。 b 、 f エリアRMS。 c 、 g 平均身長。 d 、 h 最大z -範囲

DMEM + FBSで乾燥したRE-NPのAFMおよび光学画像。 a – d オプティカル( a )およびAFM画像( b – d )5μmの自己組織化構造を示すDMEM + FBSの。 e – h CCD( e )およびAFM画像( f – h )PrF 3 の 500nmの樹状突起自己組織化構造を示すDMEM + FBS培地のNP。 i – l CCD( i )およびさまざまな倍率のAFM画像( j – l )のLaF 3 100nmの樹状突起自己組織化構造を示すDMEM + FBSのNP
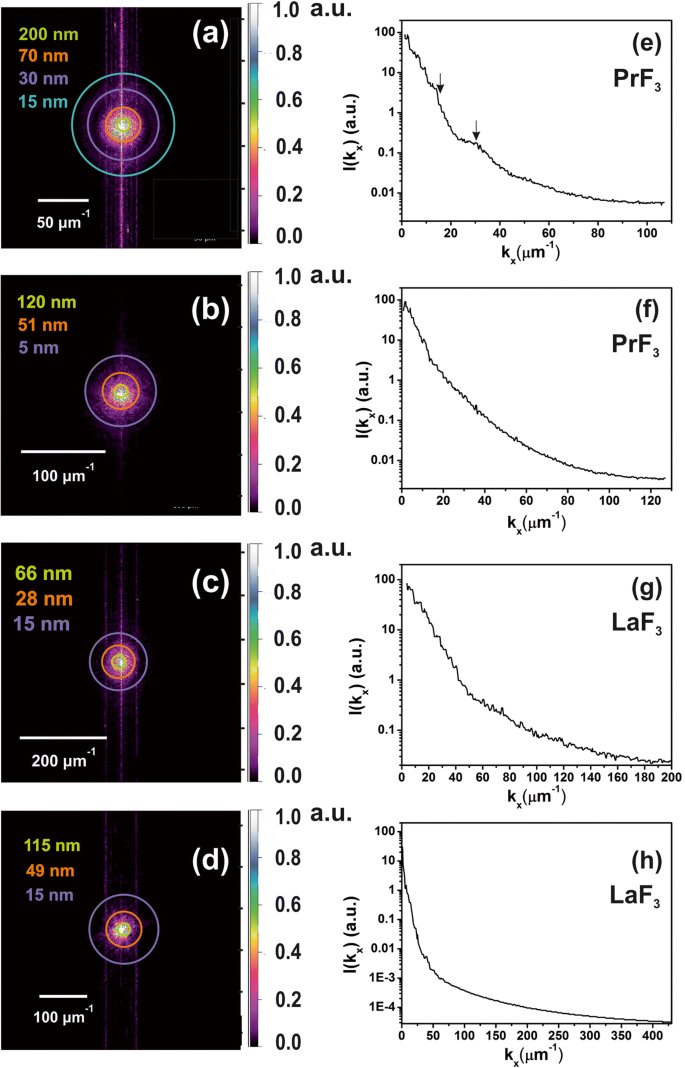
z の2D-FFTスペクトル -液体懸濁液中の小さなサイズのNPの存在を証明するDMEM + FBS培地中の乾燥RE-NPの高さ分布。 a 、 b PrF 3 のz高さ分布 AFM画像からのNP、図2(a1、b1)。小さい z -高さの特徴(〜5 nm)は( b で識別されました )。 c 、 d LaF 3 のz高さ分布 AFM画像からのNP、図2(c1、d1)。 e 、 f z のパワースペクトル -PrF 3 の高さ波数ベクトル x に沿ったNP -軸。 g 、 h z のパワースペクトル -LaF 3 の高さ波数ベクトル x に沿ったNP -軸
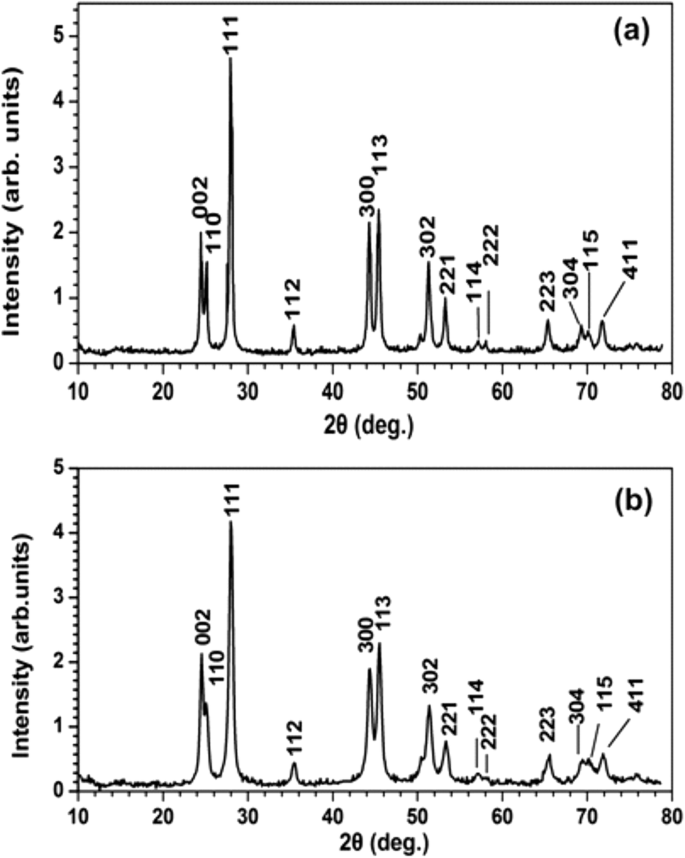
a のXRDスペクトル PrF 3 および b LaF 3 NP。 PrF 3 の平均表面積直径 およびLaF 3 NPはそれぞれ〜23±4および〜15±4nmでした
DLS
濁った不均一な混合物は、細胞培養培地にRE-NPを添加することによって最初に形成され、液体懸濁液中のRE-NP間の相互作用の複雑さを証明しました。両方のPrF 3 について、RE-NPの小さな(<10 nm)、小さい(> 10および<20nm)、および大きいサイズ(> 20 nm)の構造が特定されました。 およびLaF 3 可溶物(図1a–d)。
大型RE-NP(PrF 3 の場合は55〜83 nm)の平均流体力学的半径(MHR)値 LaF 3 の場合は99〜296 nm )は、NPの濃度レベル(0.1〜10 kg m -3 )に直接および反比例して追跡されました。 )ウシ胎児血清(DMEM + FBS)を含む液体ダルベッコ改変イーグル培地で、図1g、h。また、小型LaF 3 のMHR およびPrF 3 NPは一定のままで、RE-NPのさまざまな濃度レベルで、それぞれ10.66±0.74nmと10.64±0.40nmでした。 RE-NPのMHRは、少なくとも6日間変化しませんでした。 RE-NP懸濁液を乾燥させた後、強い相互作用によって安定化された大きなサイズの凝集体が沈殿を強制したため、RE粉末を再溶解することは不可能でした。
RE-NPのAFMおよびTEMイメージングと表面分析
0.1 kg m -3 でのDMEM + FBSにおける小型RE-NPの信頼できるサイズ分布と統計 AFM(スキャン領域1×1および2×2μm 2 )およびTEMイメージングも適用されました(図2(a1–d1)および図3(a1、b1))。 DMEM + FBS中のRE-NPの液滴をガラス基板に移した後、NPの平均サイズと平均フェレット直径の両方から比較的多数の非凝集の小さなRE-NPが同定されました[38]。図2(a2–d2、a3–d3)および図3(a2、b2、a3、b3))。また、AFMおよびTEMフェレット直径の角度分布のヒストグラム(NPのより大きな寸法の場合)は、両方のRE-NPが±(44–62 o の間の2つの方向に沿って優先的に配向されていることを示しました。 )、 x を基準に -軸(図2(a4–d4)および図3(a4、b4))。
それにもかかわらず、 z -NPの高さ分布は、全体的なNPサイズ分布に関する直接的な情報を提供しませんでした。これは、( x の最初の推定のための便利な比較ツールです。 、 y )z-heightと( x 、 y )ディストリビューションは相互に関連したままです[38]。
両方のPrF 3 の平均表面パラメータ およびLaF 3 さまざまなAFMスキャン領域の乾燥懸濁液の様子も図4に示されています。小さな z -高さの値は非常に均一な z を示しました -小さい1×1μm 2 の両方のRE-NPの高さ分布 エリアをスキャンします。それどころか、RE-NPとより大きなスキャン領域の両方で、 z -高さの分布はかなり広かった。低い z -小さなスキャン領域での高さ分布値は、液体懸濁液中の小さなサイズのRE-NPの存在を反映しています。 DMEM + FBS培地の表面パラメーター値は平均してエタノールよりも大きく、図5の多面的な構造化と2D-FFTデータ(図6)と一致して、タンパク質とRE-NP間の複雑な反応状態を示しています。 )。全体として、LaF 3 NPは、乾燥懸濁液で興味深い応答を示し、PrF 3 よりも広範な表面粗さパラメーターを示しました。 NP。
FFT
2D-FFTスペクトルで選択した半径に色付きのリングを追加しました(図6a–d)。サイクルは、ユークリッド空間でのさまざまなNPサイズ分布を表しており、ピクセルのサイズ(1.9〜3.9 nm)に等しい小さなサイズから、
XRD
XRD分光法は、結晶構造を特徴づけ、PrF 3 のサイズに関する補足情報を提供しました。 およびLaF 3 NP(図7)。両方のRE-NPの標準的な円筒相構造に対応する鋭い回折ピークは、凝集相の高い結晶状態を示しています。 Scherrerの式(\(\ tau =\ frac {0.9 \ lambda} {\ beta \ cos \ left(\ theta \ right)} \ Big)\)を使用すると、平均平均円の面積(MEAC)τ PrF 3 の およびLaF 3 NPはそれぞれ〜23±4および〜15±4nmと推定されました。
VUV分光法
ハイドロスコピックPrF 3 のVUV透過スペクトル CaF 2 に堆積したNP層 125 nm(〜10 eV)から190 nm(〜6.5 eV)までの基板を図8に示します。以前はPr 3+ の遷移に起因していた140〜170nmのVUVピーク 地面からの3価イオン 4f 4f5d のStarkコンポーネントへの電子状態構成 YF 3 内の電子配置 、LaF 3 、KY 3 F 10 およびLiLuF 4 単結晶マトリックスであり、それらは水VUV吸収帯と重なり、PrF 3 に結合した水分子が存在することを示しています。 およびLaF 3 結晶。
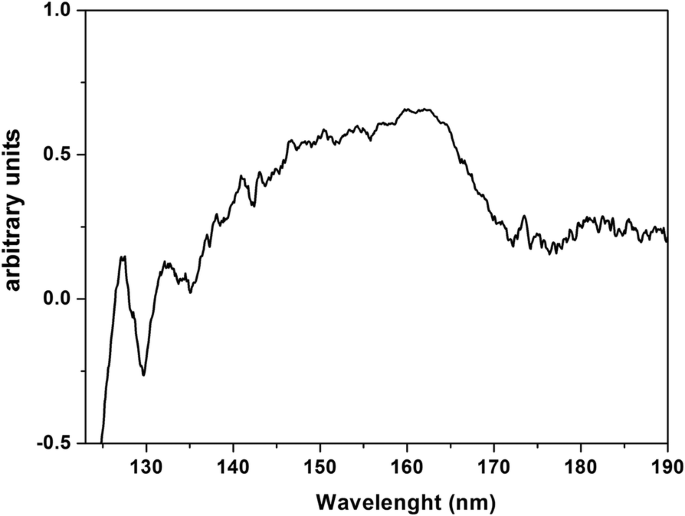
PrF 3 のVUV透過スペクトル 乾燥したCaF 2 に沈着した水懸濁液中のNP 基板。スペクトルは、PrF 3 内での水の付着とトラップを示しています NP
実行可能性テスト
サイズ分布分析とRE-NPの統計に続いて、水溶性テトラゾリウム塩(WST)生存率テストを使用して、PrF 3 の毒性を監視しました。 およびLaF 3 3つのヒト癌細胞株のNP、肺癌に由来するA549、結腸癌に由来するSW837、および乳癌に由来するMCF7。 DMEM + FBS(A549、SW837)およびウシ胎児血清(RPMI + FBS)(MCF7)を含むRoswell Park Memorial Institute培地中の3つの異なる濃度のRE-NP懸濁液(0.5、1、および5 mM)を使用しました。細胞株を最初に96ウェルプレートに置き、一晩付着させた。細胞増殖の線形領域内にあり、翌日の飽和を回避するために(図9a)、PrF 3 を含む新鮮な培地 およびLaF 3 懸濁液を添加し、RE-NPを添加してから24時間後と48時間後、または細胞プレーティングの最初の瞬間から48時間後と72時間後に生存率試験を実施しました。ただし、3つの濃度と3つの培養細胞株では、培地を交換せず、培養に追加のRE-NPを追加しなかった場合、過成長の違いが検出されました。これにより、実験の初期条件が変更されます。また、〜5×10 4 未満の細胞濃度をプレートすることは不可能でした。 3つの細胞株のコンフルエンスが小さすぎて測定可能な細胞増殖を保証できないため、ウェルあたりの細胞数。最適な実験設定は〜5×10 4 に設定されました ウェルあたりの細胞数。
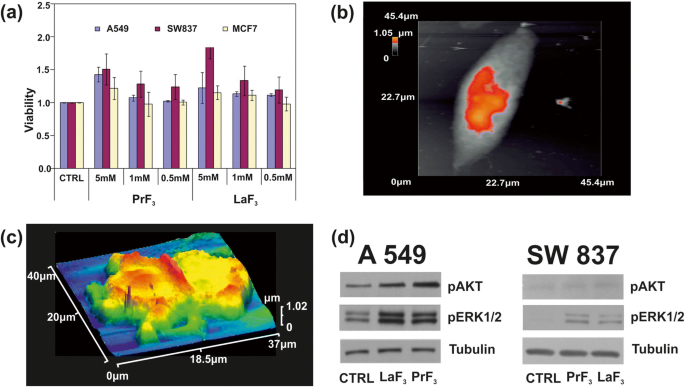
a 異なる濃度のPrF 3 で処理された3つの異なる癌細胞株(A549、SW837、MCF7)のWST生存率アッセイのヒストグラム およびLaF 3 生物学的媒体中のNP。 b 単一のA549がん細胞のAFM画像。 c DMEM + FMSのRE-NPで分裂したA549癌細胞のAFM画像。 d AKTおよびERK経路を持つA549、SW837細胞のWbリン酸化分析
より高い濃度(5 mM)では、両方のRE懸濁液について、すべての細胞株で上昇成長が得られました(図9b、c)。その中で最も高い成長値はSW837ラインでした(86%、LaF 3 )。あまり目立たないが、それでも関連性がある、5 mMのMCF7細胞株では細胞の異常増殖(15%)が認められた。 t テスト統計分析( p とフィッシャー F 腫瘍細胞の生存率の値)は、腫瘍細胞の成長が24時間で不飽和であることを示しました。それは、RE-NPの実行可能性と集中を結び付ける未知の物理法則に従った(追加ファイル1)。
リン酸化アッセイ
2つのタンパク質のリン酸化状態もテストされました(図9d)。 A549およびSW837細胞株で特定の抗体とWbアッセイを使用して、5mMのLaF 3 を含むDMEM + FBSで増殖させました。 およびPrF 3 対照細胞(CTRL)と比較して、24時間のNP、処理細胞におけるERK1 / 2およびAKTの高いリン酸化活性が得られました。
ディスカッション
癌細胞の相対的増殖速度は、両方のRE-NPのより高い濃度レベルで上昇していました(図9a)。ただし、DMEM + FBSのRE-NPのMHR値は直接追跡されました(PrF 3 )および反比例(LaF 3 )RE-NPの濃度を0.1〜10 kg m -3 (図1g、h)。したがって、平均サイズが約55 nmを超えるRE-NPは細胞増殖に影響を与えないはずであり、腫瘍増殖に実際に関与できるのは小さなサイズのNPのみです。
RE-NPのサイズと構造
小さなRE-NPの識別
実験データから、RE-NPの平均サイズ、分布、および統計パラメータが抽出されました。 t を適用する 乾燥したPrF 3 のNPの「平均等面積円」の「帰無仮説」の検定統計量 およびDMEM + FBS懸濁液、 p ランダムに選択された2つのAFM画像間のNPの直径の値は〜0.001でした(追加ファイル2)。 MEAC直径(63 nm)の値は、AFMデータから自信を持って抽出され、これはDLSデータからのMHR値と同等でした(図1g)。
逆に、ランダムに選択されたLaF 3 からのMEAC直径の値 サンプルは、平均MEAC直径が26 nmで、棄却確率値が高いことを示しました( p =0.07)、LaF 3 の発散する振る舞いを指します 液体懸濁液で。 MEACの直径とDLS(296 nm)からのMHRの不一致(図1)は、LaF 3 での相互作用の複雑さによるものです。 再びサスペンション。実際、2×2μm 2 の場合 AFMチップスキャンエリア、平均 z -高さは〜140 nmで、大きなサイズのLaF 3 の存在を示しています。 基板上の液体懸濁液から移動したNP(図4)。 「ランダムに選択されたTEMサンプルからの等しいMEAC直径値」の「帰無仮説」の場合、 p 値も小さかった( p =0.001)。両方のRE-NPについて、両方のPrF 3 のTEMデータとXRDデータの組み合わせから抽出された平均MEAC直径値 およびLaF 3 高い p を示した 値、 p =0.29と0.06であるため、TEMデータとXRDデータの間に相関関係はありません。 TEM、AFMのみ(PrF 3 )およびDLSデータは、MEACの直径とコアシェルの値を抽出するのに十分な信頼性がありました(追加ファイル2)。
また、フェレット直径の非等方性角度分布は、両方のPrF 3 およびLaF 3 異方性の角度分布はナノ結晶間の強い電気極性相互作用を示しているため、構造は高度に分極した誘電体でした。 LaF 3 の多様な分極状態 懸濁液中の凝集状態の相対効率を縮小し、乾燥サンプルの表面粗さパラメーターを拡大する役割を果たしました。
5μLおよび0.1kg m -3 の濃度のランダムAFM画像のソフトウェア粒子分析 サイズが15nmおよび10nm未満の〜22および〜11のRE-NPの数を特定しました( p =0.001)、およびTEM画像からの〜60 RE-NPの数( p =0.001)〜4μm 2 の領域 、したがって、懸濁液中に小さなサイズのRE-NPが存在することを確認し(図3(c)、追加ファイル2)、DLSでは検出されませんでした。
RE-NPの構造と形状
RE-NPのサイズ分布は、エタノール懸濁液とDMEM + FBS懸濁液で異なります(図4)。多様性は、吸着されたタンパク質、炭水化物、電解質、およびRE-NPの表面間の異なる分子相互作用を所有しており、RE-NPと細胞との特定の相互作用を調節する非常に複雑な有機マント(コロナ)の形成につながります。 DMEM + FBS培地。
吸湿性RE-NPとDMEM + FBSにトラップされた水分子間の相互作用も、コアシェル形成に不可欠でした。それはまた、調製の初期段階での相互作用の間のタンパク質およびコンフォメーション変化に大きな影響を及ぼしました。 NPの表面対バルク比が懸濁液中で高い値を示すにつれて、タンパク質を吸収する能力を含むRE-NPの有効な安定性と物理化学的、機械的および流動特性は非常に変化しました[39,40,41]。 。
AFMおよびTEMによる液体(DLS)および固化懸濁液中のRE-NPのサイズ分布の比較は、RE-NPが有機形状の内部にカプセル化され、タンパク質性シェルがREコアを囲むコアシェル誘電体構造を形成することを示しました。ガラス基板上に堆積したDMEM + FBS懸濁液中の固化したRE-NPから画像化されたAFMも、多面的なRE-NPとタンパク質コロナ複合体の形成を示しています(図5)。乾燥した媒体は結晶構造の規則的な自己組織化パターンを形成しましたが(図5a–d)、乾燥したRE-NP懸濁液は、AFMのデジタルカメラでも見ることができるいくつかの黒い斑点を持つアモルファス層状構造を示しました(図5e–l) 。より高い光学倍率では、DMEM + FBSの両方のRE-NPで、樹状突起タイプの構造とともに、培地のみの場合よりも小さい球状の離散凝集体も検出されました。どちらも相互作用の複雑さを示しており、表面パラメータの結果(図4)。最高のAFM解像度(1×1μm 2 )でも エリア)、図5の最後のレーンでは、両方のRE-NPの乾燥構造で、分解能の限界が約5nm以内の孤立したRE-NP凝集体は確認されませんでした。光学画像に示されている、長さ1〜2μmの黒色の球状菌糸体は、コアシェルRE-NPの大きな凝集層でした。 RE-NPとDMEM + FBSの間の反応の複雑さは、純粋なDMEM + FBSの長期的な自己組織化細長い構造から樹状突起構造への変換によって視覚化されました。
結果は、タンパク質シェル内にカプセル化された単一のRE-NPコア構造の図を示しています。これらの構造は、有機物と電解質に囲まれており、どちらもRE-NPと交差反応したため、検出できませんでした。 PrF 3 のVUVスペクトル は、140〜170 nmのスペクトルピークを示しています(図8)。イオン遷移は、最大で145から180nmに拡張されたVUV吸水帯と重なります。 168nmで。 4f6s のスペクトルシグネチャのみ maxima を使用した電子構成 132および127nmでスペクトルに存在しました。ただし、これらのバンドは、高吸湿性のPrF 3 に水の存在を証明する可能性があります。 サスペンション。水は、122 nmを中心とするVUVスペクトル範囲に豊富な構造化された吸収帯を持ち、コアシェルNPに水分子が存在することを示しています。
メカノセンサーのアクティブ化
外力によるインテグリンの活性化
TSRの3D構造的性質に加えて、RE-NPによる発癌経路の活性化[24]は、細胞の生存能力を維持するためのいくつかの自然進化の原理に基づいています。まず、LABSで特定の外部リガンドに結合すると、TSRスペクトル全体に沿ったコンフォメーション変化が一連のカスケード経路を強調し、腫瘍細胞の成長を引き起こします(図9c、d)。信号の伝達は、さまざまなタンパク質鎖を介して原形質膜を通って進みます。シグナル伝達は、高親和性の外力に応答するインテグリンのコンフォメーション変換を介して行われました(図10a、b)。
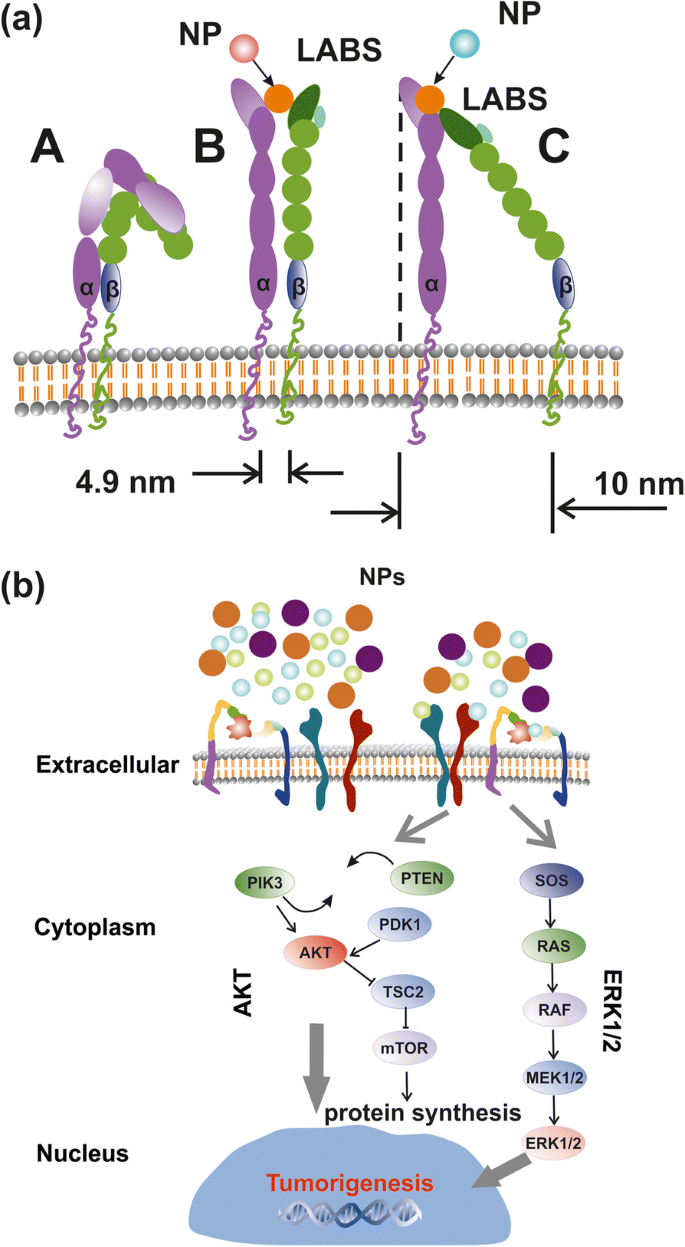
NPおよびシグナルトランザクション経路によるインテグリン活性化の簡略化されたレイアウト。 a 低(A)、中(B)、および高親和性強度(C)でのインテグリンの構造と立体配座の形状。 b AKTおよびERK1 / 2は、外部インテグリン刺激を介してRE-NPによって活性化されるトランザクション経路をシグナル伝達します
ランダムな「ノイズ」によって癌細胞の増殖を防ぐ「生命の持続可能性」と「生存法」のため、外力の強さは制限された値の範囲内にある必要があり、また外力刺激は癌細胞内の多数のメカノセンサーの長期間。癌を刺激する外部強度は、通常の条件下での原子間分子力の強度よりもわずかに大きくなければなりません。室温での配位子の熱エネルギー( kT =0.025eV、 T =298K)、分子結合の通常の熱応力が〜0.05 nmの場合、LABSに作用する平均熱力は1.2×10 -12 に留まります。 N.原則として、〜10×10 −12 を超える力 細胞上のメカノセンサーのセット全体にコヒーレントに作用するNは、腫瘍細胞のシグナル伝達を活性化するはずです。その結果、ECMの通常の状態での熱的および機械的ストレスを無視すると、LABSとNP間の電気的極性相互作用を介したインテグリンの活性化は、癌細胞でのシグナル伝達を開始し、腫瘍形成を開始する可能性があります。
インテグリンの構造と形状
直立したコンフォメーション状態のインテグリン受容体は、細胞膜から上向きに約20 nm伸びています[42](図10a)。リガンド結合ポケットの近くのヘッドピースにあるものを除いて、2つのαサブユニットとβサブユニットの間に接触がない場合、αサブユニットとβサブユニットは十分に分離されており、細胞質尾部は最大8nmまで伸びています[42]。 。 αサブユニットとβサブユニットで囲まれた円錐図法(傾斜高さ20 nm、円形ベースの直径5〜10 nm)は、細胞膜の表面に〜19〜〜80 nm
メカノセンサーとRE-NPの相互作用
ERK½およびAKTアクティベーション
TEM画像とF、La、Prの元素マッピングは、RE-NPが細胞内に浸透できなかったことを示しています。それらはA549細胞膜の周りに集まり(図11)、TSRの活性化により外力が細胞増殖を刺激できることを確認しました[44]。 Pr原子は、細胞膜の境界の周りに分布していました。細胞内の少数のF、La、Prの同定は、RE-NPのエンドサイトーシスとは関連していませんでしたが、細胞の赤道周期における2つの細胞半球の投影からのRE-NPの画像でした。

A549細胞の表面境界でのRE-NPのTEM画像と元素分析。 a 小さいサイズのLaF 3 のTEM画像 A549細胞を取り巻くNP。 b セル周辺に分布するRE-NP中のF原子の元素分析。 c La原子の元素分析。低濃度のLa原子は、X線の散乱効率がかなり小さいことに関連していました。 d – f ( a と同じ – c )PrF 3 の場合 RE-NP
また、両方のRE-NPがAKTリン酸化を増強できることも明らかであり、特にA549細胞では(図9d)、SW837細胞株のAKT経路活性の定常状態レベルが高かった。 MCF7細胞株のリン酸化レベルは検出限界を下回り、比較的低レベルの増殖と一致していました。 ERK1 / 2 [36]およびAKTの高いリン酸化レベルが、A549およびSW837細胞株で検出されました。適切なサイズのNPが細胞のメカノセンサーと相互作用して、細胞増殖を開始するための正しい力を提供すると、細胞増殖が開始されました[45、46]。 ERKおよびAKT経路は、TSRによって刺激されたり、さまざまなマイトジェンリガンドと選択的に結合したり、機械感覚グループの活性化によって刺激されたりするため、細胞外スプリングを介していくつかの癌細胞タイプで頻繁に活性化されました。相互作用は、細胞の表現型に従って、癌細胞を制御不能で無限の成長に追いやる継続的な細胞内刺激の原因でした。生存率テストも48時間と72時間実行されましたが、すべての細胞株の増殖は、細胞プレーティングの最初の瞬間から48時間と72時間で飽和しました。
細胞とイオンの相互作用
同様に、フッ化物アニオンは最も反応性の高い電気陰性元素であり、スクリーニングされていない 4f の平均半径の広がりです。 LaおよびPrの3価イオンの電子配置は比較的大きく、電気双極子相互作用を介して高い電気表面電荷が発生する可能性があります[47]。
重要な問題の1つは、特定の部位に結合する単一のイオンが腫瘍細胞の成長を活性化できるかどうかということです。 4f の投影面積が 単一のREイオンの電子配置は S 4f =0.040および0.043nm 2 ( 4f の近似球面幾何学の場合 電子配置と 4f 平均軌道半径〜 r 4 f =PrおよびLaイオンの場合はそれぞれ0.113および0.117nm)、細胞膜の全領域にわたる単一REイオンまたは他の同等サイズのイオンの一般的な上限数は〜S c でした。 / S 4f = N 4f 〜7.9×10 9 REイオン;腫瘍細胞上のインテグリンの平均数の上限を少なくとも2桁上回っている数。細胞の相対的な異常増殖は濃度の上昇とともに上昇していたため(図9a)、腫瘍細胞の増殖がリガンド部位への単一三価REイオンの特異的結合[48]によって引き起こされる可能性は低い[49,50,51]。実際、多数のREイオンが細胞の成長を飽和させているはずであり、したがって細胞の生存率はREイオンの濃度に依存しないままであるはずです。
インテグリンとRE-NPの相互作用
数pNの必要な力の範囲内で、NPからのインテグリンの効率的な活性化のために、NPとLABSの間の相互作用は、細胞のインテグリンの大部分を長期間活性化するはずです。細胞増殖に最も好まれるケースでは、NPの数は細胞表面のインテグリンの数と同じままである必要があり、LABSとNPの間の相互作用力は、一定の(長期的な)作用を得るために魅力的でなければなりません。 。腫瘍細胞を取り巻く球形NPの薄い球殻は、体積を占めていました\({V} _ {sc} \ approx 4 \ uppi {R} _c ^ 2x \)、ここで R c =5μmはセルの半径であり、 x ≈20nmは、隣接するインテグリン受容体と V の間の分離距離の半分です。 sc ≈6.3x10 9 nm 3 。各インテグリン受容体が1つのNPとのみ相互作用するという要件を正当化するために、細胞上のインテグリンのセット全体に対する上記の要件を満たすためのNPのサイズの最初の推定値は、球殻の体積を除算することによって得られます V sc インテグリンの数で。細胞半径5μmの簡単な計算は、球殻体積 V 内のインテグリンのセット全体を活性化するNPの半径の限界を示しています。 sc ≈6.3x10 9 nm 3 セルを覆うことは、ボリューム V を分割することによって得られます sc インテグリンの数\({\ overline {N}} _ {int} \ approx 2 \ \ mathrm {x} \ {10} ^ 5 \)および n int ≈1.6x10 7 。球状NPの体積は3.15×10 4 を表します および3.93×10 2 nm 3 それぞれ。したがって、インテグリンと相互作用するNPの半径は、約20〜5nmの間にあります。インテグリンの数の1桁の変動を考慮して、\({\ overline {N}} _ {int} \)、インテグリンと相互作用するNPの半径はそれぞれ〜27〜〜3nmです。
同様の簡単な計算を適用し、RE-NPの濃度レベルの実験的限界内(0.1〜10 kg m -3 ) )、最大 PrF 3 の数 MHR 55〜83nmおよびLaF 3 MHR 296〜100 nm NP(図1g、h)が腫瘍細胞の表面を覆っている V sc 4.1×10 4 の略 –2.1×10 4 および17.1×10 2 –1.5×10 4 NP。これらの値は、細胞表面のインテグリンの数よりもかなり下に配置されます。 PrF 3 の濃度が上昇する場合 およびLaF 3 0.1および10kg m -3 から 、PrF 3 の数 およびLaF 3 懸濁液中のNPは、NPのサイズが降順または昇順のいずれかで上昇する必要があります。癌細胞の生存率はより高い濃度レベルで上昇するため、現在の実験構成では、55〜296nmのサイズのRE-NPが癌細胞の有糸分裂の原因となる可能性は低いです。
また、DLSデータから、10〜20 nmの両方のRE-NPのサイズは、さまざまなRE濃度で一定(10.6 nm)のままでした。細胞表面を覆うこのサイズのRE-NPの数は、3.7×10 5 の間です。 および1.5×10 6 。この数は、細胞表面のインテグリンの平均数\({\ overline {N}} _ {int} \ approx 2 \ \ mathrm {x} \ {10} ^ 5 \)に匹敵します。したがって、実験的観察と一致して、小さなサイズのRE-NPのみが、細胞表面上のすべてのインテグリンを刺激することによって癌細胞の成長を刺激する能力を持っています(図1g、h、および9a)。
セル表面( S )上のMEAC直径(TEM)が2〜10および10〜15nmの小さなサイズのRE-NPの数 c =314μm 2 )は1.3×10 4 の略です および1.8×10 4 それぞれRE-NP。これらの値は\({\ overline {N}} _ {int} \ approx 2 \ \ mathrm {x} \ {10} ^ 5 \)より1桁低いままであったため、小さなサイズのRE-NPにも次のような効力がありました。濃度とともに生存率の値が上昇するという実験結果を正当化します(図9a)。また、腫瘍細胞の粗い表面(図9c)は空洞を形成することができ、そこで小さなサイズのRE-NPがトラップされ、細胞のメカノセンサーがトリガーされます。最も重要なのは、電気双極子相互作用を介してインテグリン受容体を活性化する能力を持っているのは、小さなサイズのRE-NPだけです(下記参照)。
EGFRとRE-NPの相互作用
EGFRを介して細胞の異常増殖を刺激できる小さなサイズのNPの上限は以前は14nmに設定されていましたが[52]、EGFRを刺激するNPの現実的なサイズは<5 nm [53]である必要があります(図12)。腫瘍細胞の表面のEGFRの面積数密度は〜1.4×10 -4 を表します nm −2 表面上のEGFRの総数 S c セルの数は〜4.2 x 10 4 の間に留まります および10 5 [54,55,56]。 5〜10 nmのサイズのRE-NPは、34 NPの数に留まりました(図3)。この数を細胞の表面に外挿する S c 、RE-NPの総数は〜10 4 のままでした NP、A549細胞上のEGFR受容体の数と一致する数。したがって、EGFRには、多数の小さなサイズのRE-NPによっても相乗的に活性化される可能性があります。
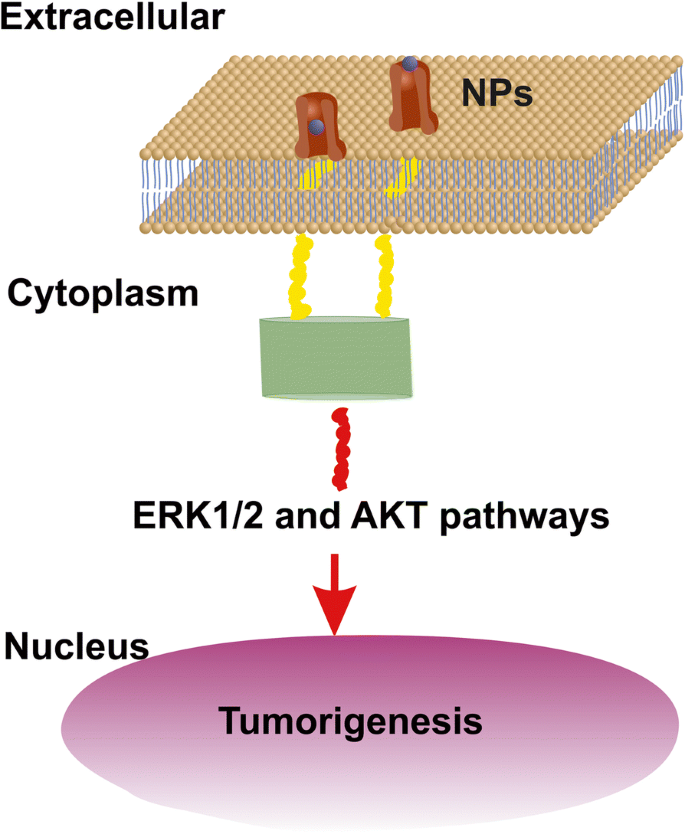
AKTおよびERK1 / 2は、EGFR刺激を介してRE-NPによって活性化されるシグナル伝達経路を示します。 EGFRは小さなサイズ〜5 nmNPによってのみ活性化されます
RE-NPとLABS間の電気双極子相互作用
上記の実験結果は、電気双極子相互作用を介した小さなサイズのコアシェルRE-NPによるLABS刺激による癌細胞増殖の仮説によって裏付けられています。
実際、コアシェルRE-NPからLABSに作用する平均電気双極子力\(\ left \ langle {\ overrightarrow {F}} _ {V_2} \ right \ rangle \)には、2つの項が含まれます(図13dおよび付録、式A22)。最初の放射状の項は、距離 r を分離する4乗に反比例します。 1 RE-NPとLABSの間であり、NPのサイズにも比例します。 2番目の極項は、分離距離 r の両方に反比例します。 1 とNPのサイズの二乗
$$ \ left \ langle {\ overrightarrow {F}} _ {V_2} \ right \ rangle =-\ frac {G_1 {N} _2 {N} _1 \ d {e} ^ 2} {4 {\ varepsilon} _0 \ {r} _1 \} \ theta \ left(\ 3G \ frac {b} {r_1 ^ 3} {\ widehat {r}} _ 1 + \ frac {\ theta} {2 {b} ^ 2} {\ widehat { \ theta}} _ 1 \ right)\ kern0.75em(1)$$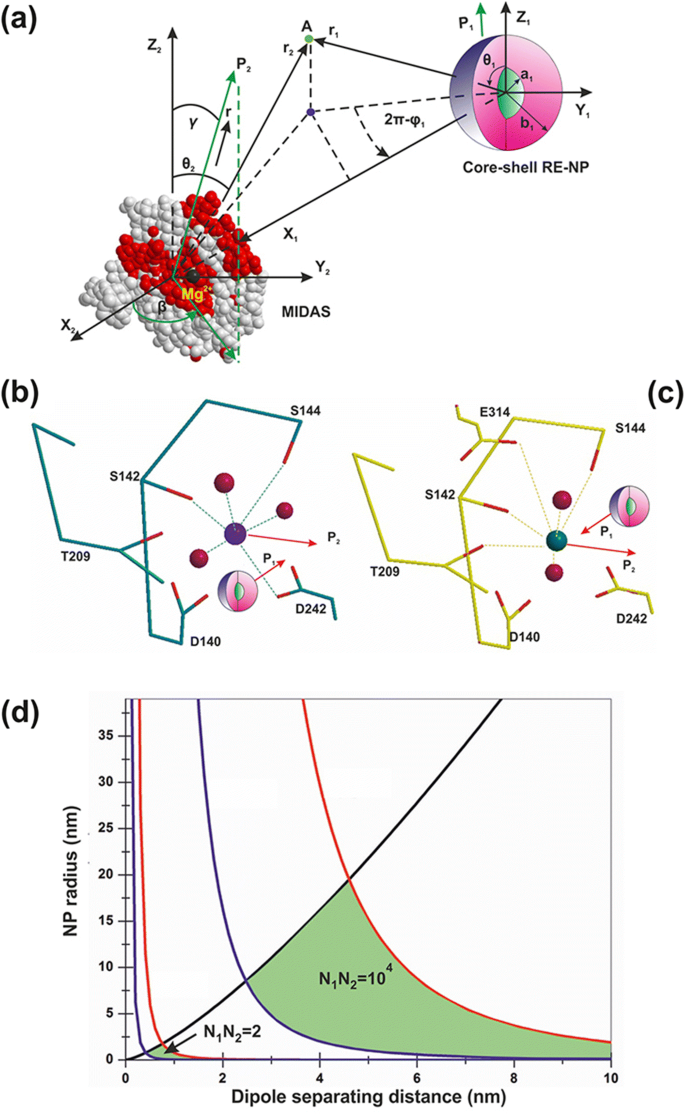
a 1つのコアシェルRE-NPと1つのLABS間の電気双極子相互作用。 b 、 c MIDASの近くのREコアシェルNP( b )およびADMIDAS( c )接着部位。 d RE-NPのサイズの軌跡領域(緑)と、2つの充電状態でのLABSとコアシェルRE-NP間の分離距離
式で。 1、 G および G 1 はNPの幾何学的要因であり、コアシェルまたはコア球状構造のいずれかを記述しています。付録、式。 A6およびA14; N 1 、 N 2 RE-NPおよびLABS表面の表面電子の数です。 d および b LABSの原子軌道の有効な特徴的な空間拡張、〜0.1 nm、およびRE-NPの半径です。 e およびε 0 は電子電荷と真空誘電率および\(\ theta =\ frac {d} {r_1} <0.01 \ rad \)です。 RE-NPのコアは結晶性半導体材料であるため、固有の多数の表面およびボリュームの欠陥サイトが、電子がコアボリューム内で自由に移動できるようにする高密度の疑似電子エネルギーレベルの原因でした[46]。 。その結果、コアシェル構造は高度に分極される可能性がありました。したがって、LABSは、近接した分離距離での電気双極子相互作用を介して、コアシェルRE-NPによって効率的にアクティブ化できます。コア核の高い分極効率は、2つの異なる方向に沿ったNPの選択的配向によって実験的に確認されました(図2(a4–d4)および図3(a4、b4))。
極性相互作用力は、幾何学的係数 G にも比例します。 1 、付録、式。 A14。培地の誘電率、シェル構成、およびREコアコンポーネントの一般的な値は、εを表します。 1 =78、ε 2 =10およびε 3 =15。コアシェルとコア半径の比率が b / a の場合 1から50の範囲内で、幾何学的係数 G 、 G 1 ほぼ一定の値を保持します( G =0.2、 G 1 =0.01)そしてそれらは両方の球形コア( b / a )で自己同一です =1)および球形のコアシェル。 LABSドメインを形成する開いたまたは閉じたa-I-MIDASドメインの永続的または誘導された分極は、Mn 2+ を持つ6つの配位した水酸素原子軌道に起源があります。 またはMg 2+ 球形の幾何学的構成に配置されたイオン[7](図12a–c)。
式の電気双極子力として。 1は、放射状(第1項)と極性成分(第2項)のベクトル和を表します。ただし、最後の項が最初の項よりも優先されます。
$$ {r} _1> \ sqrt [3] {6G} b \ sim b \ kern0.75em(2)$$この場合、LABSは、すべての(b / a)比で極力成分からアクティブ化されます。最も重要なのは、この項がNPのサイズの2乗に反比例することです。これは、わずかまたは小さいサイズのLaF 3 NPは癌細胞の増殖を活性化しました。
さまざまな r の一般的な極力項 1 および b 値とさまざまなΝ 1 、Ν 2 LABS / MIDASをアクティブにする充電状態は制限内にとどまります[57,58,59,60]。
$$ {10} ^ {-12} N <\ frac {G_1 {N} _2 {N} _1 \ d {e} ^ 2} {8 {\ varepsilon} _0 \ {r} _1 {b} ^ 2 \ } {\ theta} ^ 2 <{10} ^ {-9} N \ kern1em(3)$$不等式3は、サイズ b に関連しています REコアシェルNPの分離距離 r 1 束縛電子または自由電子の数Ν 2 、Ν 1 2つの双極子の表面に。ポイントの軌跡( r 1 、 b )異なる表面電荷状態の不等式3を満たすΝ 1 、Ν 2 黒、赤、青の線で囲まれています(図13c)。 RE-NPのタイプには特定の仮定がなかったため、結果はどのタイプの分極化NPにも等しく適用できます。
表面電子数の代数積 N 1 および N 2 LABSとRE-NPの(バインド済みまたは無料)は、それぞれ N でした 1 N 2 =2、RE-NPのサイズとインテグリン活性化の分離距離の軌跡は<1nmでした。より高い充電状態では、 N 1 N 2 =10 4 、軌跡領域は、より広いRE-NPサイズにまたがり、分離距離領域の値のセットは、それぞれ0.5 nm〜19 nmから2.5〜15nmになります。
上記の分析から、特定の分離距離 r でLABSをアクティブにできるのは、小さいサイズまたは小さいサイズのNPのみであることがわかります。 1 電気双極子相互作用の強さは、NPのサイズの2乗に反比例して減衰します。図13cから、 N の充電状態の場合 1 N 2 =5 x 10 4 、LABSをアクティブ化できるNPのサイズは制限によって制限されます
$$ 2.5 \ \ mathrm {nm} 最も重要なのは、図13cから、充電状態が高くなると、軌跡領域(緑色の領域)とRE-NPのサイズの両方が増加することです。結論
癌は複雑な病気です。腫瘍は非常に不均一であり、細胞増殖は、他の要因の中でも、細胞と絶えず変化する細胞外マトリックスとの間の動的相互作用に依存しています。ランダムなゲノム変異、細胞内のシグナル伝達に加えて、細胞増殖の活性化は、細胞外細胞マトリックスの微小環境と膜のメカノセンサーとの間の機械的、熱力学的および電気的極性相互作用によって引き起こされます。ここでは、3つの異なるヒト癌細胞株(A549、SW837、MCF7)における腫瘍細胞増殖が、EGFRの同期的かつ相乗的な活性化によって、または小さなサイズのRE-NPとLABS間の電気的双極子相互作用を介して活性化される可能性があることを示しました細胞上のインテグリンの。
インテグリン活性化の前提条件となる力は10 -12 の間にあるはずだからです および10 -9 N、細胞増殖を引き起こすアクティブなRE-NPのサイズは、特定の制限内にある必要があります。癌の活性化は、LABSとNPの両方の電気的表面電荷とそれらの分離距離によって指定されます。この電気双極子活性化力は、NPの半径の反比例二乗則に従い、実験結果と一致して、小さなまたは小さいサイズのRE-NPのみが電気双極子相互作用を介して癌細胞の成長を刺激する能力を持っていることを証明します。
メソッド
RE-NPの合成
PrF 3 NPは共沈によって合成されました。簡単に説明すると、4gのPr 2 O 3 ポリプロピレンガラスビーカー内の110mLの10%硝酸に、3gのNaFと一緒に攪拌しながら添加しました。混合物を50°Cに加熱し、透明な薄緑色の溶液が現れるまで45分間撹拌した。それからそれはろ過されました。 25%の水和アンモニウムを加えることにより、混合物のpHを4に調整した。次に、混合物を再び20分間撹拌した。最後に、沈殿したNPを遠心分離により蒸留水で洗浄しました。
LaF 3 NPは、La 2 の混合物に同じプロトコルを適用することによっても合成されました。 O 3 (4 g)およびNaF(3 g)。両方の調製物から、NPを含む懸濁液のアリコートを構造解析のために風乾し、残りの部分を生物学的研究のために水懸濁液として保管しました。
NPの懸濁液は、完全なDMEM + FBS細胞培養培地で、水に懸濁したNPを培地に直接添加して最終濃度を5mMにすることにより調製しました。次に、5 mMのストック溶液から始めて、溶媒としてDMEMを使用するいくつかの後続の希釈液を、それぞれ1mMと0.5mMの最終NP濃度に調製しました。
RE-NPのサイズ分布
XRD
PrF 3 の結晶構造とサイズ およびLaF 3 NPは、X線分光法によって特徴づけられました。X線回折計(島津XRD-7000S)は、グラファイト単色化Cu-Ka放射線(1.5406Å)を使用して、10°から80°の2θ範囲にあります。統計では、すべてのピークのτの加重平均が使用されました。 β以外の重み付けでは、XRDスペクトルのすべてのピークの相対強度が考慮されました。対応するエラーには、読み取りエラー(0.3 mrad)と平均の標準エラー( se )が組み込まれています。 =σ /√Ν 。
DLS
水およびDMEM + FBS懸濁液中のRE-NPのサイズ分布とMHRは、マルチアングル動的および静的光散乱装置(PHOTOCOR-FC)を使用して632.8nmのDLSと37°Cの直角で比較するために決定されました。 MHR(ストークス半径)の値とNPのサイズ分布は、自己相関スペクトルとDynaLSソフトウェアとのストークス-アインシュタインの関係から計算されました。純粋なDMEM + FBSでの散乱光の強度はRE-NP添加剤の場合よりも20倍低かったため、純粋なDMEM + FBSでの凝集タンパク質のレベルは、DMEM + FBS培地でのRE-NPの混合懸濁液と比較して無視できました。 MHRおよびRE-NPのサイズ分布とサイズ誤差は、DLS機器からのデータをDynaLSソフトウェアでフィッティングおよび処理することによって得られました。これにより、MHRを10〜10 の主要なサイズ分布のさまざまなスペクトルドメインで計算できます。 2 〜10 2 –10 3 nm、追加ファイル2。
AFM
15 nm未満のサイズ分布はDLSの下限範囲に近いため、AFMを適用して小さなサイズ分布を評価しました。液体懸濁液中のRE-NPの濃度が低く、ガラス基板上の液滴の乾燥速度が遅い場合、堆積物は液体懸濁液のサイズ分布を反映していました[37]。 RE-NPをエタノールまたはDMEM + FBSに分散させた後、マイクロピペットを使用して懸濁液をきれいなガラス基板上に滴下し、AFMイメージングおよび分析(diInnova、Bruker)のために室温で空気中で乾燥させました。 AFMは、リン(n)をドープしたシリコンカンチレバー(Bruker、RTESPA-CP)を使用し、公称ばね定数が40 nN / nmで、共振周波数が300 kHzの周囲条件で、タッピングモードで実行されました。さまざまなサイズの表面積(0.5×0.5–50×50μm 2 )は、0.2 Hzのスキャンレートで高空間分解能(512px×512px)で画像化され、「スキャンエリアフィルタリング」[37]を介して異なるサイズ分布のドメインを識別しました。 SPM LabAnalysis V7ソフトウェアによる形態素解析から、粒子のサイズ分布、形状、および凝集段階が決定されました。
さまざまなスキャン領域のNPのサイズは、粒子分析クロマチックバー(MEAC直径に基づくFiji統合ROIカラーコーダー)でも顕著でした(図2(a1–d1))。 AFM画像は、適切な z を使用してバイナリ画像に変換されました。 -高さのしきい値。処理された画像のすべてのピクセルには、 z だけでなく情報が含まれていました -ピクセル領域の高さだけでなく、ピクセル領域に粒子が存在する場合。 x -MEACとフェレットの直径のヒストグラム(図2(a2–d2、a3–d3))は、「Image J 1.51n Fiji配布ソフトウェア」を使用して、正しい z で抽出されました。 -高さのしきい値。 PrF 3 の場合、ピクセルあたりのサイズ解像度は3.9および1.9nmでした。 およびLaF 3 それぞれ。
粒子同定、ノイズ抽出、粒子面積データは、Fijiソフトウェアの「ParticleAnalyzer機能」で処理しました(図2(a1–d1))。粒子径のヒストグラムも分析されました。等面積の円の直径(図2(a2–d2))とフェレットの直径または「キャリパーの直径」(最大)の両方 粒子の全方向の直径)(図2(a3–d3))、その方向はフェレット角(図2(a4–d4))であることが分析されました。同定されたすべての粒子を考慮して、平均等面積円直径および平均フェレット直径を計算した。関連するエラーには、すべてのAFM画像の実際のピクセルサイズと平均の標準エラー( se =σ /√Ν 。
t テストは、ランダムに選択された図の間のすべてのAFM画像で平均粒子径が同じであるという「帰無仮説」に基づいてAFM画像のすべてのセットに対して実行されました(図2(a1–b1、c1、d1))。 p 値( t に基づく帰無仮説の確率 配布は無効です)は、追加ファイル2に示されています。
TEM
TEMイメージングで上記のパラメータを計算するために同じ手法が採用されました(図3(a1–b4))。原子分解能TEM(Hitachi HT7700 Exalens)は、グルタルアルデヒドで固定されたA549細胞上の細胞外または細胞内RE-NPの付着を画像化しました。 F、La、Prの元素分析も実施しました(Oxford Instruments X-Max80T)。
2D-FFT
( x のNPサイズ分布に関する追加情報 、 y )平面は、関係を使用してNPのAFM画像の2次元フーリエ変換からも抽出されました
$$ I \ left({k} _x、{k} _y \ \ right)=\ iint f \ left(x、y \ right)\ exp \ left(i {k} _xx \ right)\ exp \ left( i {k} _yy \ right)dxdy $$ここで f ( x 、 y )は、ある点( x、y )でのサイズ関数です。 )、 k x 、 k y は、同じ点と I にある逆ユークレディアン空間の関連する波数ベクトルです。 ( k x 、 k y )は、関数 f の「スペクトル密度」です。 ( x 、 y ) k の時点で x 、 k y 。ほとんどのアプリケーションでは、 f ( x 、 y )は z です -ポイントでのNPの高さ( x、y )および z = f ( x 、 y 。
For a set of discrete data, such as the digitised AFM images, the 2D-FFT was used instead of 2D Fourier transform in the continuous space. For a m × n X-matrix (pixels of an AFM image), the 2D-FFT transform takes the form
$$ \kern1em {Y}_{p+1,q+1}=\sum \limits_{j=0}^{m-1}\sum \limits_{k=0}^{n-1}{\omega}_m^{jp}{\omega}_n^{kq}{X}_{j+1,k+1\kern1.25em } $$where \( {\omega}_m^{jp}={e}^{2 pi/m},{\omega}_n^{kq}={e}^{2 pi/n} \) are the associated frequencies. Then, an appropriate shift along the y -axis was performed and the integers m, n, p, q, k were translated into lengths and inverse lengths respectively by a multiplication with the pixel’s size of the image.
Water Trapping in RE-NPs
VUV Spectroscopy
To appraise the state of water in RE-NP’s complexes during the initial stage of suspension preparation, the adsorption of water molecules on the surface of the hygroscopic PrF3 NPs was identified with a laboratory-made VUV (110–180 nm) absorption spectrometer. It consists of a hydrogen lamp operating in a longitudinal stabilised discharge mode at 10 kV, a stainless steel vacuum chamber and a VUV monochromator (Acton VM502), equipped with a solar blind photomultiplier (Thorn EMI 9412 CsTe) and a laboratory-made data collection system. Thin layers of PrF3 NPs suspensions in water were prepared and dried on 1-mm-thick VUV-grade CaF2 substrates by applying the “drop-casting method”. Then, the CaF2 substrates were placed in the optical path between the hydrogen lamp and the VUV monochromator in a vacuum. The stainless steel 316 vacuum chamber was evacuated initially to 10 − 7 mbar using two turbomolecular pumps at a differential pumping configuration (Edwards EXT 100/200, pumping speed 150 ls −1 )。 However, a high outgassing rate of PrF3 sets an upper limit to the background pressure in the vacuum chamber ~ 8.5 × 10 −5 mbar。 The relatively low background pressure of both compounds irreversibly damages the VUV optics and the turbomolecular pump after few hours of operation and therefore it sets certain experimental constraints, preventing an equivalent registration of LaF3 spectrum because of high outgassing rates and a low background operating pressure (< 10 −4 mbar). The experimental data (light transmitted through the sample film on CaF2 window) were fitted to a logarithmic response for calculating the transmittance.
Cell Culture and Growth Assay
Cell Growth
The A549 and SW837 cell lines were maintained in DMEM+FBS, whereas the MCF7 lines were in RPMI+FBS. Both media supplemented with 10% fetal bovine serum (FBS), 1 × penicillin, 1 × streptomycin and 2 mM l-glutamine. Cells were incubated at 37 °C, 5% CO2 in a humidified atmosphere.
The WST viability test was used to monitor the intrinsic toxicity of PrF3 およびLaF 3 NPs for three human cancer cell lines, A549, SW837 and MCF7. For the viability assay, three different concentrations of RE solubles (0.5, 1 and 5 mM) in DMEM+FBS (A549, SW837) and RPMI+FBS (MCF7) were used. The initial number of cells seeded in the 96-well plates was ~ 5 × 10 4 細胞/ウェル。 This amount of cells was plated 24 h prior to the RE-NPs treatment of cells in order to allow enough time for the cells to attach properly to the plate (wells) and to attain the optimum growing conditions. Subsequently, the viability test was performed 24 h after RE-NPs addition, or 48 h after the initial cell cultures were placed in the wells. As we did not observe any cell reduction, but on the contrary cell-overgrowth, especially with the SW620 cell line at 5 mM, the cell confluence quickly reached 80–90% of its initial value after 24 h of the addition of RE-NPs or 48 h from the initial plating.
Five microliters of WST solution was added to each well and the plate was incubated for 1 h during the growth state. The absorbance at 450 nm of each well was measured using a microplate reader (Biorad, x Mark). Each experimental point for each cell line and each RE suspension was extracted from two samples and triplicated every 2 days (total of 108 samples).
F test was used for every set of cell viability measurements. Here, the “null hypothesis” was that the relative to the CTRL “mean viability value was the same at different concentrations within the same cell line”. With this null hypothesis, an unknown law connecting tumour cell viability and RE-NPs concentration was identified. The p value (probability the null hypothesis to be rejected) was also tested from the F distribution Additional file 1.
Western Blotting and Antibodies
Total proteins were extracted with 60 μL of radioimmunoprecipitation assay (RIPA) lysis buffer (20 mM Tris-HCl (pH 7.5); 150 mM NaCl, 1 mM Na2 EDTA; 1 mM EGTA; 1% NP-40; 1% sodium deoxycholate; 2.5 mM sodium pyrophosphate; supplemented with proteases inhibitors 1 mM β-glycerophosphate; 1 mM Na3 VO4 1 μg/ml; leupeptin) and the Wb assay was performed according to standard protocols (Fig. 9b). Briefly, total proteins (50 μg) were separated by SDS-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to nitrocellulose membrane. Blots were incubated overnight at 4 °C with appropriate primary antibodies. The antibodies used were tubulin code sc-8035, from Santa Cruz (final concentration 1:1000 in blocking buffer); p-ERK (E-4) code sc-7383, from Santa Cruz (final concentration 1:500 in blocking buffer); and p-AKT (Thr308) code 9275S, from Cell Signaling (final concentration 1:1000 in blocking buffer).
Wb bands are collected from different blots showing quality control of antibodies specificity. Numbers at the top of the phosphorylation images show grey scale levels from 0 (black) to 168 (grey) (maximum value ), indicating activation at a non-saturated mode.
略語
- 2D-FFT:
-
Two-dimensional fast Fourier transform
- ADMIDAS:
-
Adjacent MIDAS
- AFM:
-
原子間力顕微鏡
- AKT:
-
Protein kinase B
- CTRL:
-
Control cells
- DLS:
-
動的光散乱
- DMEM:
-
ダルベッコの改良イーグル培地
- ECM:
-
Cell-extracellular matrix
- EGFR:
-
Epidermal growth factor receptors
- ERK:
-
Extracellular signal-regulated kinase
- F.A.:
-
Feret angle
- F.D.:
-
Feret area diameters
- FBS:
-
ウシ胎児血清
- LABS:
-
Ligand adhesion binding site
- MEAC:
-
Mean equal area circle
- MHR:
-
Mean hydrodynamic radius
- MIDAS:
-
Metal ion-dependent adhesion sites
- NGFR:
-
Nerve growth factor receptor
- NP:
-
ナノ粒子
- RE-NPs:
-
Rare-earth nanoparticles
- RIPA:
-
Radioimmunoprecipitation assay
- RMS:
-
Root mean square
- RPMI:
-
Roswell Park Memorial Institute medium
- SDS-PAGE:
-
Sodium dodecyl sulfate-polyacrylamide gel electrophoresis
- SyMBS:
-
Synergistic metal ion binding sites
- TEM:
-
透過型電子顕微鏡
- TSR:
-
Transmembrane signal receptors
- VEGFR:
-
血管内皮増殖因子
- VUV:
-
Vacuum ultraviolet
- Wb:
-
Western blot assays
- WST:
-
Water-soluble tetrazolium salts
- XRD:
-
X線回折
ナノマテリアル
- 半導体ナノ粒子について
- 生体適合性FePO4ナノ粒子:ドラッグデリバリー、RNA安定化、および機能的活性
- HT29およびSPEV細胞株に対するAuナノ粒子の影響のinvitro研究
- 炭化水素中の金ナノ粒子のサイズ制御と安定化のための分散剤としての修飾ハイパーブランチポリグリセロール
- 6-メルカプトプリンとニューロン透過性ペプチドで修飾された金ナノ粒子によるSH-SY5Y細胞増殖の促進
- マゲマイトナノ粒子はナノザイムとして機能し、セイヨウアブラナの成長と非生物的ストレス耐性を改善します
- 細胞イメージング用の近赤外発光Cr3 + / Eu3 +共ドープ亜鉛ガロゲルマネート持続性発光ナノ粒子
- 抗菌剤として銀ナノ粒子で装飾された酸化グラフェンベースのナノコンポジット
- 丸い形の金ナノ粒子:シロイヌナズナの根の成長に対する粒子サイズと濃度の影響
- 反応性高周波(RF)熱プラズマによるバイメタルタングステン銅ナノ粒子のその場合成
- ポリ(γ-グルタミン酸)は、Fe-Pdナノ粒子によるp-クロロフェノールの脱塩素化の促進を促進します