


環境変化に応じた細菌のマイクロコンパートメントシェルタンパク質の自己組織化の安定性と変動性
要約
細菌のマイクロコンパートメント(BMC)は、原核生物界に広まっているタンパク質性の自己組織化オルガネラです。多面体シェルを使用して主要な代謝酵素と経路をセグメント化することにより、BMCは炭素同化、病因、および微生物生態学において重要な役割を果たします。 BMCシェルは、自己組織化して定義されたアーキテクチャを形成する複数のタンパク質ホモログで構成されています。新しいナノバイオリアクターと分子足場を開発するためにBMCを設計することに大きな関心が寄せられています。ここでは、高速原子間力顕微鏡(HS-AFM)を使用して、さまざまなpHと塩の条件下でのBMCシェルタンパク質の形成と自己組織化ダイナミクスの定量的特性評価を報告します。 400 mMの塩濃度は、シェルヘキサマーによって形成されるより大きな単層シェルパッチをもたらす傾向があり、中性pHでヘキサマー自己組織化のより高い動的速度が観察されたことを示します。また、六量体アセンブリから繊維状配列までのシェルタンパク質の変動性を視覚化します。この研究は、微小環境の変化に応じたBMCタンパク質の自己組織化の安定性と変動性に関する知識を前進させ、自己組織化と構造的堅牢性を再構築する能力を備えた合成BMC構造の合理的な設計と構築に情報を提供します。また、バイオテクノロジーアプリケーションにおけるBMCベースのナノ構造の自己組織化と形成を定量的に評価するための強力なツールボックスも提供します。
はじめに
細菌のマイクロコンパートメント(BMC)はタンパク質性の細胞小器官であり、ウイルスのキャプシドに構造的に似ており、細菌の細胞質を分割します[1]。それらは細菌の門に広く行き渡っており[2]、真核生物に見られる膜結合細胞小器官がない場合に細菌が重要な代謝経路を区画化することを可能にします[3、4]。 BMCは、管腔酵素コアをカプセル化する半透性タンパク質シェルによって形成されます。シェルは、BMC-H(Pfam00936ドメインを含む)、BMC-T(2つのPfam00936ドメインを含む)、およびBMC-P(Pfam03319ドメインを含む)を含む3種類の構造タンパク質コンポーネントで構成されています[5,6,7、 8,9]。シェルの主要コンポーネントはBMC-Hです。これは、凸面と凹面のヘキサマーとして表示され、凹面が外側を向くようにシェルファセットを並べて表示します[10](図1)。 BMC-Pは二十面体形状の頂点をキャップするために提案された五量体を形成し、BMC-Tはおそらく殻の透過性に関与する殻のファセットに位置する疑似六量体を形成します。
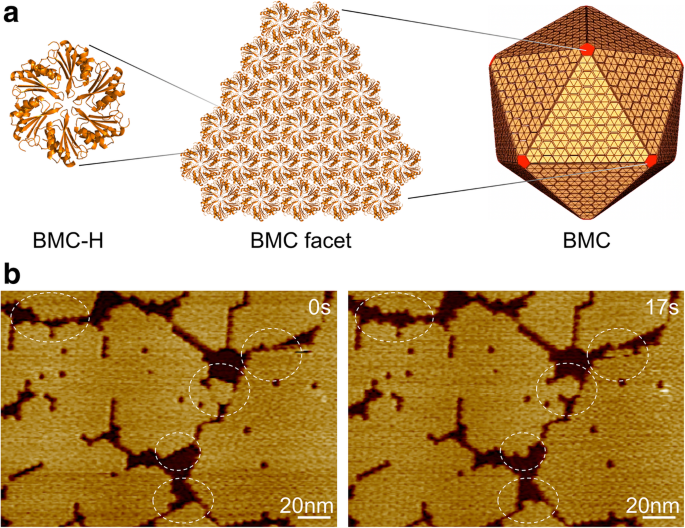
細菌のマイクロコンパートメント、シェル組織、および自己組織化。 a BMCシェルタンパク質ホモログの何百ものコピーが自己集合して二十面体タンパク質オルガネラを形成します。黄色のBMC-Hタンパク質はファセットを形成し、赤色のBMC-Pタンパク質は頂点を占めます。 b Hoch_5815BMC-Hヘキサマーで構成されるシェルファセットのAFMトポグラフ。 HS-AFMを使用して数秒以内に動的イベント(円)が観察されました
特定のタンパク質間相互作用により、BMCタンパク質の自己組織化が保証され、代謝機能を実現するための高度に定義されたアーキテクチャが形成されます。シェルタンパク質間の横方向の相互作用は、二十面体シェルの自己組織化特性を決定するための主要な要因であると考えられています[10]。 BMC-Hホモログは、2次元シート[11、12]、ナノチューブ[13、14、15、16、17]、フィラメント構造[15、18、19、20]などのさまざまな形状を形成できることが観察されています。 。
BMCは、天然に存在する細胞小器官の自己組織化、選択的透過性、および酵素カプセル化特性に基づいて、代謝酵素を封入することによるナノスケールバイオリアクターのバイオインスパイアード構築や新しい分子足場の生成など、バイオエンジニアリングにおいて大きな可能性を秘めた理想的なシステムと見なされています。新しい機能[21、22、23、24、25、26]。ただし、BMCバイオエンジニアリングでは、BMC構造の安定性や、自己組織化とBMCタンパク質凝集体の形成を効果的に操作および評価する方法など、いくつかの重要な問題に取り組む必要があります。 BMCシェルとBMC全体の構造と組み立ての調査は、X線結晶学、電子顕微鏡(EM)、蛍光顕微鏡、および動的光散乱(DSL)を使用して実施されています[10、11、16、22、27、28 、29、30、31]。最近、高速AFM(HS-AFM)を利用して、BMC-Hタンパク質の動的な自己組織化プロセスの最初の可視化を実施しました[12]。
この作業では、HS-AFMを使用して、さまざまなpHおよびイオン条件下でのBMC-Hパッチの構造ダイナミクスを監視します。これにより、BMCシェルタンパク質アセンブリの変調に関する洞察が得られ、分子分解能での定量的評価のための強力なツールが提供されます。 、BMCシェルタンパク質の自己組織化の安定性と変動性について。
メソッド
サンプル準備
Haliangium ocraceum から精製されたBMC-Hタンパク質(Hoch_5815) カーフェルド博士(ローレンスバークレー国立研究所)から親切に提供されました。バッファー交換の場合、サンプルを〜80 mg mL -1 でストックします。 Trisバッファー(50 mM Tris-HCl、pH 7.8、100 mM KCl、10 mM MgCl 2 )を0.5 mg mL -1 に希釈しました AFMイメージングの前に目的のバッファーを使用します(追加ファイル1:図S1)。コントロールバッファーは、50 mM Tris-HCl(pH 7.8)および10 mM MgCl 2 です。 。
原子間力顕微鏡
マイカおよびAFMイメージングでのサンプル吸収には、必要なバッファーを使用しました。雲母に5分間吸収した後、Hoch_5815を目的のバッファーでリンスして固定化タンパク質を除去し、AFMを使用して画像化しました(追加ファイル1:図S1)。 HS-AFM画像は、ULTRASpeed2.8μmスキャナーとUltra-ShortCantilever USC-0.3 MHzプローブ(NanoWorld)を備えたJPK NanoWizard ULTRA speed AFMを使用して、ACモードの溶液で30または40Hzでキャプチャされました。タンパク質集合の妨害を減らすために、AFMイメージング中に約100ピコニュートンの最小負荷力が適用されました[12、32、33、34、35、36]。
画像処理と分析
画像解析は、最初にJPK SPMデータ処理(JPK)を使用して実行されました。 HS-AFM画像分析は、前述のように[12]、Image SXM(http://www.ImageSXM.org.uk)でカスタムマクロを使用して実行されました。 Hoch_5815パッチのサイズを分析するために、30 Hzのスキャンレートでキャプチャされた512×512ピクセルの画像を平坦化して、XY傾斜とZしきい値を削除し、続いてタンパク質を表示するかタンパク質を表示しないようにバイナリ変換しました。粒子分析を使用して、これらのバイナリ画像のタンパク質の表面積を計算しました。パッチは、個々のパッチと隣接するパッチを識別するために、> 3ピクセル(〜2 nm)で区切られたオブジェクトとして定義されました。初期テストでは、多数のピクセルが設定されている場合、隣接するパッチは単一の連続パッチとしてカウントされる可能性があり、小さいピクセル数を使用すると、パッチ内の個々のヘキサマー間のギャップがパッチ間の境界として誤ってカウントされる可能性があることが示されました。タンパク質のダイナミクスを分析するために、40 Hzのスキャン速度でキャプチャされた256×256ピクセルの一連の画像を分析し、フレームあたり約6.4秒の時間分解能を実現しました。バイナリ画像は、違いのAFM画像を示すために、シリーズの前の画像から差し引かれました。差分画像の粒子分析を使用して、組み立てられたタンパク質と分解されたタンパク質の面積を数えた。動的速度の計算に使用される式は次のとおりです。
$$ \ mathrm {Rate} \ \ mathrm {of} \ \ mathrm {dynamic} \ \ mathrm {events} \(R)=\ frac {\ mathrm {Number} \ \ mathrm {of} \ \ mathrm {hexamers} \ \ mathrm {added} \ \ mathrm {or} \ \ mathrm {removed} \ \ mathrm {in} \ \ mathrm {a} \ \ mathrm {s} \ mathrm {eries} \ \ mathrm {of} \ \ mathrm {frame} \ mathrm {s} \ kern0.5em(N)} {\ mathrm {Total} \ \ mathrm {surface} \ \ mathrm {area} \ \ mathrm {of} \ \ mathrm {protein} \ \ mathrm { in} \ \ mathrm {frame} \ kern0.5em(A)\ times \ mathrm {time} \ kern0.5em(T)}、$$ここで N しきい値処理された差分画像の白と黒のピクセルの合計を、そのスケールの単一のヘキサマーに対応するピクセル数で割ったものを表します(追加ファイル1:図S3、図S5)。データは平均±標準偏差(SD)として表されます。統計分析は、指定された多変量分散分析または二元配置分散分析を使用して実行されました。
結果
粘液細菌 Haliangium ocraceum のBMC-Hタンパク質(Hoch_5815)を使用しました 、 Escherichia coli で発現した そして、6回対称の六量体として特徴付けられます[12]。 Hoch_5815ヘキサマーは、自己組織化して2番目のタイムスケールで単層シートを形成できます。これは、二十面体BMCアーキテクチャの基本的な構造コンポーネントを表しています(図1a)。 HS-AFMイメージングにより、シートフラグメントの動的アセンブリと組織の柔軟性を視覚化し(図1b)、開発されたイメージング分析を使用してBMC-Hタンパク質のパッチサイズと動的速度を定量的に推定できます(「方法」セクションを参照)。
pH変動への応答
Hoch_5815の自己組織化能力の全体的な指標として、パッチサイズの変化を測定しました。パッチサイズは、pHが3から10に上昇するにつれて増加し(図2a、追加ファイル1:図S2、表S1)、低pH条件よりも高pHの方がHoch_5815タンパク質の自己組織化に適していることを示しています。これは、pH 6で不溶性であり、pH 8でナノチューブの規則正しい配列を形成し、pH10で分解する傾向があるRmmHタンパク質の集合挙動とは多少異なります[13]。さらに、HOCH_5815の自己組織化の高度な構造的変動が観察されました(図2a、追加ファイル1:図S2の大きなSDで示されています)。
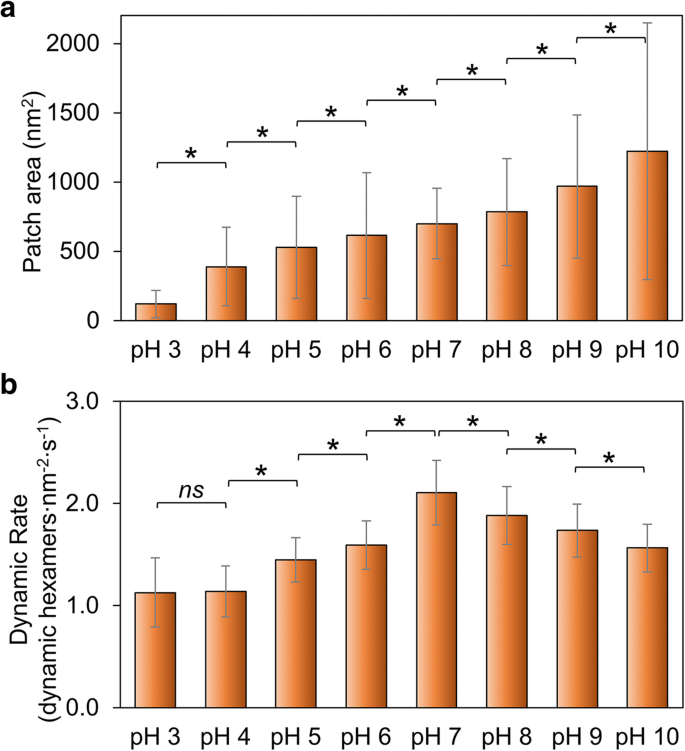
Hoch_5815の自己組織化に対する環境pHの影響。 a AFMによって決定されたHoch_5815の個々のパッチの平均表面積( n =50)(追加ファイル1:図S2)。 b HS-AFMによって決定された動的イベントの平均レート( n =50)。 * p <0.05、 ns 有意ではない(多変量分散分析)
シェルシートにおけるHoch_5815タンパク質の自己組織化に関するAFMイメージングにより、シェルシートの形成はヘキサマーの組織化と分解の組み合わせに起因することが明らかになりました[12]。さらに、Hoch_5815のタンパク質間相互作用の安定性を調べるために、さまざまなpHでのHoch_5815の自己組織化ダイナミクスと動的イベントの割合を調べました(追加ファイル1:表S2)。自己組織化のダイナミクスの速度は、pH 7で最も高く、酸性条件とアルカリ性条件の両方で低下します(図2b、追加ファイル1:図S3)。特に、酸性条件、特にpH7からpH6で急速に低下し、図2bに示すように、pH4とpH3の間で比較的一定に見えます。
pHは、ヘキサマー-ヘキサマー界面にあるアミノ酸残基の静電特性に大きな影響を与える可能性があります。酸性条件で観察されたダイナミクスの減少とシェルパッチのサイズの減少は、Hoch_5815の自己組織化能力が低下していることを示しています。アルカリ性条件で観察されたダイナミクスの減少とシェルパッチのサイズの拡大は、安定したヘキサマー-ヘキサマー相互作用を示唆しますが、Hoch_5815ヘキサマーのダイナミクスの増加は、中性pH条件での柔軟なヘキサマー-ヘキサマー相互作用を意味します。
塩濃度の変動への対応
また、バッファーの塩濃度がHoch_5815の組み立てに影響を与えるかどうかを確認しました。低濃度(100〜200 mM)のMgCl 2 、CaCl 2 、およびKCl、Hoch_5815タンパク質は、高濃度(300〜500 mM)で組み立てられたものよりも比較的小さなパッチを形成します(図3a;追加ファイル1:図S4)。 500 mMで、2層または多層のHoch_5815シートが観察されました(追加ファイル1:図S4)。これらの観察結果は、より高いイオン強度が、炭素同化のためのカルボキシソームのシェルタンパク質であるCcmKによるより広範囲で秩序だった2D結晶の形成を促進する可能性があるという以前の発見と一致しています[37]。ただし、NaCl濃度が50から500 mMに増加すると、RmmHによって形成された高度に秩序化されたナノチューブが分解され[13]、シェルヘキサマーによるフラットシートと管状形状の形成を仲介する潜在的に異なるメカニズムを示しています。
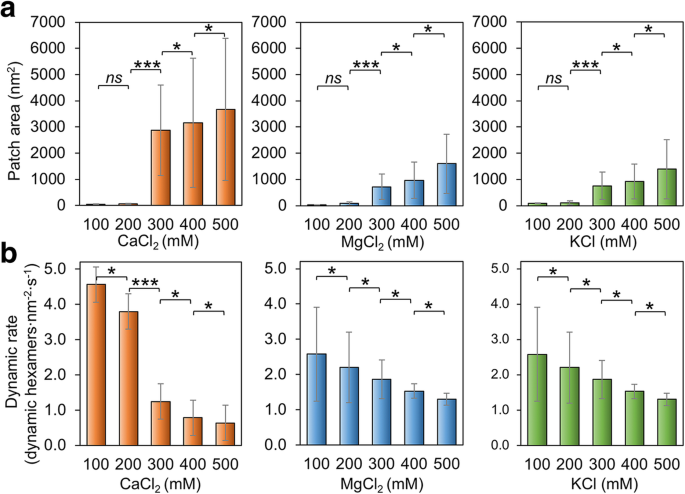
Hoch_5815の自己組織化に対する塩濃度の影響。 a 100〜500 mM CaCl 2 の範囲でAFMによって測定された平均パッチ面積 、MgCl 2 、およびKCl( n =50)。塩濃度の上昇により、パッチサイズが増加しました。 200〜300 mMの間でパッチ面積の有意な変化が観察されました(*** p <0.001、* p <0.05、 ns 重要ではない、双方向ANOVA)。 b 100〜500 mM CaCl 2 の範囲で高速AFM画像シリーズから決定された動的イベントの平均レート 、MgCl 2 、およびKCl( n =50)。塩濃度が100mM変化するたびに、動的イベントの発生率が大幅に変化しました(*** p <0.001、* p <0.05、 ns 重要ではない、双方向ANOVA)
さらに、MgCl 2 の変化によって引き起こされるHoch_5815自己組織化の変動 とKCl濃度は比較的似ています。対照的に、パッチサイズの変化は、CaCl 2 の場合に最も顕著になります(最大3000倍の増加)。 濃度が200mMから300mMに上昇し(図3a)、CaCl 2 に対するHoch_5815自己組織化の感度が高いことを示唆しています。 MgCl 2 よりも またはKCl。
Hoch_5815の自己組織化の動的速度は、緩衝塩濃度の変化によっても影響を受けます。 MgCl 2 の増加 、CaCl 2 、またはKCl濃度により、Hoch_5815動的速度が低下する可能性があります(図3b、追加ファイル1:図S5)。より高い塩濃度で観察されたパッチサイズの増加を考えると(図3a)、Hoch_5815ヘキサマー間の横方向の相互作用は高塩濃度でより安定しているように見えます。 CaCl 2 の変更 濃度はより顕著な反応を示し、動的イベントの発生率は200〜300 mMの間で大幅に変化しましたが(図3b)、MgCl 2 の変化に対する反応は とKClは比較的類似しており、パッチサイズの変更と一致しています(図3a)。興味深いことに、アセンブリイベントと分解イベントの比率が最も高いのは、400mMのMgCl 2 で観察されました。 、CaCl 2 、またはKCl(追加ファイル1:表S2)。これにより、400 mMの塩の下で大きくて安定した単層Hoch_5815アセンブリが形成されました(追加ファイル1:図S4)。 500 mMで観察された2層アセンブリも安定しており、ヘキサマーの移動速度が低くなっています。
BMC-Hタンパク質アセンブリの柔軟性
スキャン力を100pNに減らすことで、BMCタンパク質のアセンブリに対するAFMチップスキャンの影響を最小限に抑え、個々のヘキサマーの分子分解能AFM画像を取得しました(図4)。組み立てと分解の両方のイベントを同じビューで見ることができ、チップスキャンアーティファクトの代わりにBMCシェルの動的な性質が組み立てられることを確認します[12]。 HS-AFMイメージングでは、Hoch_5815タンパク質凝集の変動性も明らかになりました。わずか10mMMgCl 2 の存在下でpH7.5でサンプルをイメージングする場合 驚くべきことに、2番目のタイムスケールでHoch_5815ヘキサマーの分解に伴う繊維状構造の形成が時折観察されました(図4a)。これらの繊維のような構造は、シェルヘキサマーによって組み立てられたナノチューブの束と同様に、並列に密に詰めることができます[13、14、15、16]。ただし、2本のファイバー間のスペースは3.72±0.31 nm( n )です。 =30)、平均身長は2.46±0.22 nm( n =30)、Hoch_5815ヘキサマーによって形成されたシェルシートよりも少ない(3.45±0.16 nm、 n =25)(図4b–f)。これらのファイバー構造は、イメージング中にかなり柔軟で動的であり、さまざまなサイズのストレートまたはコイル状のアーキテクチャを表示できます。 Hoch_5815ヘキサマーの分解に伴う繊維構造の外観と、単層ヘキサマーシートと比較して高さが低いことを考えると、これらの繊維状構造は、ヘキサマーから分解された個々のHoch_5815ペプチドによって形成されていると推測されます(図4g)。 。特定のバッファー条件(低イオン強度など)での基質吸収により、Hoch_5815ペプチドのアルファヘリックス側が基質表面に付着し、ペプチドが隣接するペプチドと直線的に結合する可能性があります。ヘキサマー内の相互作用はおそらく強い[5]。シェルタンパク質凝集の変動性の根底にある詳細なメカニズムは、さらに解明されていない。
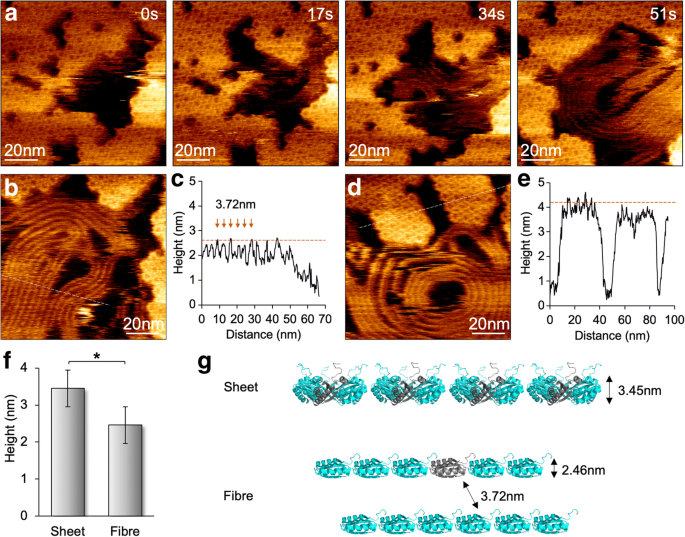
HS-AFM下のシェルシートアセンブリとともに繊維構造の形成とダイナミクス。 a AFM画像シリーズで示されている、Hoch_5815ヘキサマーで構成されたシェルシートの分解中の繊維状構造の外観。 b 繊維構造のAFMトポグラフ。 c 断面分析(パネル b の破線 )は3.72±0.31 nm( n )の間隔を示します =30)隣接する繊維構造間で、平均高さは2.46±0.22 nm( n =30)。 d Hoch_5815ヘキサマーで構成されるシェルパッチのAFMトポグラフ。 e 断面分析(パネル d の破線 )は、Hoch_5815ヘキサマーの平均高さが3.45±0.16 nm( n )であることを示しています。 =25)。 f 繊維状の構造は、Hoch_5815ヘキサマー(* p )で構成されるフラットシートと比較して高さが低くなっています。 <0.05、双方向ANOVA)。 g Hoch_5815モノマーのストリングを表す繊維状構造の提案された組織化と形成
ディスカッション
BMCは、自己組織化して高次構造を形成する何百ものタンパク質で構成されています。多数のタンパク質ホモログで構成されるBMCシェルは、タンパク質の自己組織化と相互作用を研究するための理想的なシステムです。生体膜の組織化、タンパク質の集合、および生物学的システムの生理学的役割に非常に関連する物理的相互作用を分析するための強力な手法として[32、35、38、39]、AFMは組織化と自己組織化のダイナミクスを視覚化するために利用されていますBMCシェルタンパク質とBMC構造のアーキテクチャおよび機械的特徴[12、30、31、40、41、42]。この作業は、私たちの知る限り、AFMを使用した環境変化に応じた2次元シートの形成におけるBMCシェルタンパク質の自己組織化ダイナミクスの最初の定量的決定を表しています。結果は、BMC-Hタンパク質の自己組織化に固有の変動性と環境依存性を強調しています。 EMやDSLと比較して、AFMは、分子の詳細を使用してBMCタンパク質の自己組織化の動的な作用をリアルタイムで監視する大きな可能性を示しています。
タンパク質間相互作用は、BMCシェルの形成と形成において非常に重要です[10]。タンパク質濃度は、シェル形成を促進するための重要な要因としても文書化されています[41、43]。さらに、in vitro溶解度研究は、溶液中のpHとイオン強度がBMCの構造安定性[17、27]、および2次元シートの形成におけるBMCシェルタンパク質の集合挙動に影響を与える可能性があることを示しています[37、41 ]、ナノチューブ[13、17]、およびナノケージ[28]は、ウイルスキャプシドアセンブリへの影響を彷彿とさせます[44、45]。また、pH> 10および<3または塩濃度<10mMまたは> 600 mMの場合、タンパク質の沈殿が見られ、パッチは形成されませんでした(未発表のデータ)。ここでは、アセンブリの傾向とダイナミクスがpHと塩濃度に依存することをさらに示しました。シェルタンパク質は広範囲のpHで自己組織化できますが、中性のpH環境では、組織化のダイナミクスを高めることができるようです(図2b)。 300 mM以上の濃度の陽イオンは、2次元シートの形成を促進することがわかりました。 400 mMの陽イオンは、大きくて安定した単層シートの形成に望ましいようです(図3)。これらの条件は、細菌細胞の細胞質ゾル条件と一致し、生理学的に関連しています。たとえば、最も生理学的に適切な条件下では、 EのpH。コリ サイトゾルは約7.4〜7.8 [46]であり、イオン濃度は約100〜400 mMであり、タンパク質相互作用、タンパク質-リガンド結合、シグナル伝達、膜静電ポテンシャルの維持、および膜を横切るタンパク質勾配に不可欠です[47、48]。サンプルと雲母基質間の相互作用がBMCタンパク質の自己組織化にどのように影響するかについては、さらに調査する必要がありますが、AFMイメージングは、環境変動に応じたBMCタンパク質の自己組織化の動的変化を定量的に分析する機会を提供します。
ここで説明するシェルフラグメントの形成におけるBMCタンパク質の環境依存のアセンブリダイナミクスは、BMC全体の形成におけるそれらの挙動を表す可能性があります。実際、3D BMC構造は、自然界で設計された動的に維持されている細胞小器官のように見えます。 BMCは、顕著な構造上の柔軟性と不均一性を示します。 AFMナノインデンテーションによって決定されたBMCシェル構造の機械的柔らかさ[30]と、計算シミュレーションによって明らかにされたBMCアセンブリの非平衡ダイナミクス[49]は、BMCと堅牢なウイルスアセンブリの違いを浮き彫りにしました。同様に、カルボキシソームの生合成は、光やシャペロンと相関することが解明されています[50、51]。ごく最近、CcmK3とCcmK4がヘテロヘキサマーを形成し、pH依存的にカルボキシソームシェルをキャップし、カルボキシソームシェルの透過性とCO 2 を調節する手段を提供する可能性があることが示されました。 非常に動的な微小環境での同化[52]。溶液中の環境条件がBMCタンパク質の熱力学的集合にどのように影響するかについての正確なメカニズムは、たとえば、実験的研究と計算シミュレーションの組み合わせを使用して、まだ調査されていません。
BMC構造の自己組織化を考えると、BMCのエンジニアリングや、細胞代謝、酵素カプセル化、分子送達、治療などのバイオテクノロジーアプリケーションにおける新しいBMCベースのナノバイオリアクター、分子足場、生体材料の設計に大きな関心が寄せられています。 。環境変化に応じたBMCの構造的弾力性と変動性に関する高度な知識は、異種ホスト、つまり Eで堅牢なBMCベースのナノ構造を生成するための戦略に役立つだけではありません。コリ または植物[31、53、54]だけでなく、2Dナノ材料の形成、およびBMCシェルベースのタンパク質ケージの開閉を調節するための道を開き、それによって機能調節と標的分子送達を促進します。以前に、遺伝子組み換えアプローチを使用して、シェルタンパク質の界面での特定の接触とそれらの自己組織化挙動を操作することの実現可能性を実証しました[12]。この研究は、さまざまな環境でBMCシェルの自己組織化を評価および操作するためのツールボックスを強化します。
結論
要約すると、HS-AFMを利用して、さまざまなpHおよび塩条件下でのBMCシェルタンパク質の自己組織化の定量的調査を実施しました。シェルヘキサマーのより大きな単層パッチの形成は、400 mMの塩濃度で促進されることが示され、中性pHにより、ヘキサマーの自己組織化の動的速度が高くなりました。六量体アセンブリから繊維様アレイへのシェルタンパク質の組織的移行も視覚化されました。この研究は、環境条件がBMCシェルタンパク質の組織化と自己組織化を決定する上で重要な役割を果たすことを示しました。
略語
- BMC:
-
バクテリアマイクロコンパートメント
- BMC-H:
-
バクテリアマイクロコンパートメントヘキサマー
- BMC-P:
-
バクテリアマイクロコンパートメント五量体
- BMC-T:
-
バクテリアマイクロコンパートメントトリマー
- DSL:
-
動的光散乱
- E。コリ :
-
大腸菌
- EM:
-
電子顕微鏡
- HS-AFM:
-
高速原子間力顕微鏡
ナノマテリアル
- IoTと5Gの影響の評価
- プライベートネットワークの革新:帯域幅と期間をその場で設定および変更する
- スタックカップカーボンナノファイバーの原子および電子構造を明らかにする
- ボロフェンの安定性とSTM画像に関する第一原理研究
- 蒸発誘起自己組織化と強化されたガス検知特性によるワームホールのようなメソポーラス酸化スズの容易な合成
- Ag n V(n =1–12)クラスターの構造的、電子的、および磁気的特性の調査
- ポリオール媒介プロセスによるZnOナノクリップの製造と特性評価
- ロボット、コボット、自動化:パンデミックは製造業の変化の波を推進していますか?
- 製造業のリバウンドが従業員の採用と定着率をどのように変えるか
- 産業革命から自動化、そしてその間のすべてに至るまでの製造業の環境への影響
- オートメーションとロボットは世界を変えるか?