


MicroRNA-410のアップレギュレーションまたはWnt-11のダウンレギュレーションは、骨芽細胞を増加させ、破骨細胞を減少させて、大腿骨頭の骨壊死を軽減します
要約
背景
大腿骨頭の骨壊死(ONFH)におけるマイクロRNA-410(miR-410)の機能的役割についてはほとんど知られていません。したがって、本研究の目的は、Wnt-11を標的とするmiR-410を調査して、ONFHの予防における骨形成および破骨細胞のメカニズムを調節することでした。
メソッド
15個のONFHサンプルと15個の通常のサンプルが収集されました。臨床サンプル中の大腿骨頭、骨芽細胞、破骨細胞の病理学的変化が観察されました。 ONFHのラットモデルには、agomir-miR-410、Wnt-11-siRNA、またはoe-Wnt-11が注射されました。 MiR-410; Wnt-11;骨芽細胞関連因子アルカリホスファターゼ(ALP)、骨ガンマ-カルボキシグルタミン酸タンパク質(BGLAP)、およびCollα1発現。破骨細胞関連因子である酸性ホスファターゼ5(ACP5)、カテプシンK(CTSK)、およびMMP9、ならびにBcl-2およびBaxの発現は、RT-qPCRおよびウエスタンブロット分析によってテストされました。血清中の破骨細胞機能指数NTX-1およびCTX-1とともに骨形成機能指数ALPおよびOCNをELISAによって試験した。
結果
MiR-410、ALP、BGLAP、およびCollα1は分解され、Wnt-11、ACP5、CTSK、およびMMP9は臨床サンプルのONFH組織で増強されました。 miR-410のアップレギュレーションとWnt-11のダウンレギュレーションにより、ラットの骨密度(BMD)とBV / TVが向上し、大腿骨頭、大腿骨頭、脊柱のBMDレベルが上昇し、ラットの血清カルシウムとリンのレベルも上昇しました。 、骨細胞のアポトーシスを抑制しながら、ラットにおけるOCN、ALP、BGLAP、およびCollα1の発現を上昇させ、ACP5、CTSK、NTX-1、CTX-1、およびMMP9の発現を低下させました。
結論
この研究は、miR-410のアップレギュレーションまたはWnt-11のダウンレギュレーションが骨芽細胞を増加させ、破骨細胞を減少させてONFHの発生を軽減することを示唆しました。したがって、miR-410はONFHの治療の潜在的な標的として役立つ可能性があります。
はじめに
大腿骨頭の骨壊死(ONFH)は、股関節に影響を与える最も身近な病気の1つであり、激しい痛みや関節障害を引き起こし、主に中年の人に発生します[1]。中国に812万人の患者を抱える成人ONFHの治療は、依然として外科医にとっての課題です[2]。高脂血症、自己免疫疾患、凝固障害アルコール依存症、高コルチゾン症など、多くの危険因子がONFHの発生に関連しています[3]。外科的治療には、アジュバントの有無にかかわらず、自己骨髄などのコア減圧術が含まれますが、人工股関節全置換術(THR)は、関節の保持によって治療されない老人性患者または進行性骨壊死に対して保持されます[4]。しかし、ONFHは、THRを頻繁に必要とする患者の予後を満足させるものではありません[5]。したがって、ONFHの治療のための正確な治療標的を探求する緊急の必要性があります。
マイクロRNA(miRNA)は、細胞機能と遺伝子発現の主要な調節因子であり、内皮の恒常性に多大な機能を発揮し、新しい治療法となる可能性があります[6]。ある研究では、miR-410はいくつかのヒトの有害な癌の種類で異常に発現し、子宮内膜癌、肺癌、骨髄腫、および乳癌の腫瘍阻害剤として使用できることが報告されています[7、8、9、10]。別の研究では、PTEN / AKT / mTORシグナル伝達経路を抑制することによって6-ヒドロキシドーパミンによって誘発されるパーキンソン病細胞モデルに対するmiR-410の神経保護効果があることが明らかになりました[11]。さらに、miR-410は、FKBP5およびAKTシグナル伝達経路を標的とすることにより、急性リンパ芽球性白血病の小児の悪性生物学的行動を調節することが明らかになっています[12]。ある記事では、膵臓癌におけるアンジオテンシンIIタイプ1を標的とするmiR-410の阻害効果も示されています[13]。 Wntタンパク質は、システインに富む分泌型リポ糖タンパク質の一種であり、発生と疾患に多大な機能を発揮します[14]。 Wnt-11はWntシグナル伝達経路に属し、発がんにおいて極めて重要な役割を果たす正の調節因子です[15]。子宮頸癌における扁平上皮癌抗原およびWnt11の発現の臨床的重要性が報告された研究があります[16]。さらに、TGF-β1とWnt11の相乗的シグナルが、Rhoキナーゼ-アクチン-MRTF-Aシグナル伝達によって平滑筋におけるsm-α-アクチンの発現を促進することが示されました[17]。上記の文献に基づいて、本研究の目的は、ONFHの予防におけるWnt-11を調節するmiR-410の役割を調査することであり、Wnt-11を標的とするmiR-410が骨形成および破骨細胞を調節できるという仮説が提案されています。 ONFHの予防のメカニズム。
材料と方法
倫理声明
この研究は、ヘルシンキ宣言の信条に従い、首都医科大学北京四季丹病院の倫理委員会の承認を得て実施されました。参加者は、この研究に参加するための書面によるインフォームドコンセントを提供した。すべての動物実験は、国立衛生研究所の実験動物の管理と使用に関するガイドと一致していました。プロトコルは、キャピタル医科大学の北京Shijitan病院の動物実験倫理委員会によって許可されました。
調査対象
2017年1月から2018年9月までに首都医科大学北京四季丹病院整形外科で治療を受けた合計30人の患者が選ばれた。これらの患者のうち、ONFHの15人の患者がTHR手術で治療され、患者の年齢の中央値は50.6±4.3歳、体重は57.0±5.6 kgで、男性7人と女性8人でした(ONFHグループ)。大腿骨頸部骨折の別の15人の患者はTHR手術で治療され、年齢の中央値は59.6±3.3歳、体重は50.0±5.6 kgで、男性9人と女性6人(対照群)でした。 2つのグループ間で性別、年齢、体重に顕著な違いはありませんでした( P > 0.05)これは同等でした。軟骨下壊死領域の組織は、大腿骨頭の縦線に沿ってノミで採取し、-80℃で保存しました。各グループは、病理学と分子生物学によってそれぞれ調べられました。
ヘマトキシリン-エオシン(HE)染色
サンプルを4%パラホルムアルデヒドで固定し、10%エチレンジアミン四酢酸で脱灰し、脱灰溶液を週に1回交換しました。骨サンプルの色を観察し、脱灰度を測定した。完全に脱灰した後、サンプルをパラフィンに包埋し、4μmの厚さにスライスしました。焼きたての切片をキシレンIとキシレンIIに順番に10分間浸し、脱ロウした切片を無水アルコールI、無水アルコールII、95%アルコール、80%アルコール、70%アルコールにそれぞれ2分間浸しました。次に、切片をヘマトキシリンで3分間染色し、1%塩酸アルコールで2分間分化させました。次に、切片を50%、70%、80%のアルコールに2分間ずつ浸し、エオシンに5秒間浸しました。次に、切片を95%アルコール、無水アルコールI、および無水アルコールIIに3分間浸漬し、次にキシレンIとキシレンIIに順番に5分間浸漬しました。最後に、切片を中性ガムで密封し、顕微鏡で検査しました。
アルカリホスファターゼ(ALP)染色
各サンプルで1つのセクションを選択し、60°Cで60分間焼きました。パラフィン切片をキシレンIとキシレンIIでそれぞれ15分間脱ロウし、無水アルコールI、無水アルコールII、95%エタノール、90%エタノール、80%エタノール、75%エタノールに順番に5分間浸漬しました。 3回蒸留水で2分間3回洗浄しました。 ALP染色キット(NanJing JianCheng Bioengineering Institute、Nanjing、China)で調製した基質液を切片に滴下し、基質液をサンプルで完全に覆い、37°Cで15分間光を避けて孵化しました。余分な染料溶液を廃棄し、切片を直ちに発色剤Aで5分間滴下し、3回蒸留水で30秒間洗浄しました。次に、切片を発色剤Bで30秒間染色し、試薬で30秒間対比染色しました。写真は200倍の光学顕微鏡で撮影し、骨芽細胞の数は顕微鏡用の画像解析システム(Image-Proplus 6.0)を介して計算しました。
酒石酸耐性酸性ホスファターゼ(TRAP)染色
サンプルごとに1つのセクションを選択し、60°Cで60分間焙煎しました。パラフィン切片をキシレンIとキシレンIIで15分間脱ロウし、無水アルコールI、無水アルコールII、95%エタノール、90%エタノール、80%エタノール、75%エタノールに順番に5分間浸しました。切片は、準備した固定液で30秒間固定しました。切片を染色ラックに挿入し、新たに調製したTRAP染色液(Sigma、セントルイス、ミズーリ州、米国)を含むダークボックスに入れ、染色液を切片で完全に覆った後、切片をハッチングしました。 37°Cのウォーターバスポットで1時間。次に、切片をヘマトキシリンで2分間対比染色し、乾燥させた。 TRAP染色は時間とともに減衰するため、切片は密封せずに顕微鏡で直接検査されます。写真は200倍の光学顕微鏡で撮影し、破骨細胞の数を顕微鏡用の画像解析システム(Image-Proplus 6.0)でカウントしました。
免疫組織化学的染色
脱灰後、切片をパラフィンワックスに包埋し、4μmの厚さにスライスしました。切片を従来のキシレンで脱ロウし、勾配アルコールで脱水し、3%H 2 で孵化しました。 O 2 (Sigma-Aldrich Chemical Company、St Louis、MO、USA)7°Cで30分間、次に0.01Mクエン酸バッファーで95°Cで20分間煮沸しました。切片を通常のヒツジ血清作動液で10分間ブロックし、一次抗体Wnt-11(1:100、Invitrogen、Carlsbad、カリフォルニア、米国)と4°Cで一晩混合し、二次抗体(すぐに使用できる二次抗体)キット(PV-6000)、ZSGB-Bio、北京、中国)を20分間、西洋ワサビペルオキシダーゼ標識ストレプトミセスオーバルブミン作動液(SA / HRP、Beijing ComWin Biotech Co. Ltd、北京、中国)を2分間。切片は、ジアミノベンジジン(DAB)(Sigma-Aldrich Chemical Company、米国ミズーリ州セントルイス)で現像し、ヘマトキシリン(Shanghai Bogoo Biological Technology Co.、Ltd。、中国、上海)で対比染色し、密封しました。リン酸緩衝生理食塩水(PBS)は、ネガティブコントロール(NC)として一次抗体に取って代わりました。光学顕微鏡下では、陽性細胞は細胞質内に茶色の反応物とともに位置し、強い陽性発現は暗褐色がかった黄色であり、弱い陽性発現は淡褐色がかった黄色であり、陰性発現は着色していない。細胞の染色を光学顕微鏡で観察し、各セクションで5つの高倍率視野(400×)を観察し、視野ごとに100個の細胞をカウントしました。平均値は各区間の観測結果である。陽性細胞の割合と分布に従って、以下のように決定された:陰性(-)、単一細胞染色、陽性細胞は5%未満。弱陽性(+)、散乱または小細胞塊染色、陽性細胞の数は5〜24%でした。陽性(++)、フレークまたはクラスター細胞染色、陽性細胞の数は25〜50%でした。強く陽性(+++)の拡散細胞染色では、陽性細胞の数は50%以上でした。免疫組織化学の結果は、2人が独立して二重盲検スコアで評価しました。
逆転写定量的ポリメラーゼ連鎖反応(RT-qPCR)
全RNAは、Trizol抽出キット(Invitrogen、Carlsbad、California、USA)によって組織サンプルから抽出されました。 RNAの濃度と純度を測定しました。プライマーは、タカラバイオ株式会社(大津、滋賀、日本)によって考案および配合されました(追加ファイル1:表S1)。次に、PrimeScript RTキット(Takara Biotechnology Ltd.、Dalian、China)を使用して、RNAをcDNAに逆転写しました。 SYBR®PremixExTaq TM の指示に従い、反応溶液を蛍光定量PCRに使用しました。 IIキット(Takara Biotechnology Ltd.、大連、中国)。蛍光定量PCRは、ABIPRISM®7300システム(Applied Biosystems、マサチューセッツ、米国)で実装されました。 U6はmiR-410のローディングコントロールであり、グリセルアルデヒドリン酸デヒドロゲナーゼ(GAPDH)は、Wnt-11、ALP、骨ガンマ-カルボキシグルタミン酸タンパク質(BGLAP)、Collα1、酒石酸耐性酸性ホスファターゼ5(ACP5)、カテプシンの内部パラメーターでした。 K(CTSK)、マトリックスメタロペプチダーゼ(MMP)9、Bcl-2、およびBax。標的遺伝子の相対的な転写レベルは、2 -△△Ct によって計算されました。 メソッド[18]。
ウエスタンブロット分析
総タンパク質は、大腿骨頭の組織から抽出されました。各サンプルのタンパク質濃度を測定し、サンプルの負荷を脱イオン水で調整して、サイズが一定になるようにしました。ドデシル硫酸ナトリウム分離ゲルとスペーサーゲル(10%)を調製した。氷浴と遠心分離後、同量のマイクロサンプラーを用いた電気泳動によりサンプルを分離し、ゲル上のタンパク質をニトロセルロースメンブレンに転写しました。ニトロセルロースメンブレンを5%スキムミルクパウダーで4°Cで一晩密封し、一次抗体Wnt-11(1:150、Santa Cruz Biotechnology、Santa Cruz、CA、USA)、ALP(1:100、Sigma、St .Louis、MO、USA)、BGLAP、Collα1およびMMP9(1:1000)、ACP5(1:100)、CTSK(1:500、Abcam、Cambridge、MA、USA)、およびBcl-2およびBax(1: 500、Proteintech、シカゴ、米国)そして一晩孵化した。そして、サンプルを、HRPで標識された二次抗体IgG(1:1000、Wuhan Boster Biological Technology Co.、Ltd。、湖北省、中国)で滴下し、1時間インキュベートしました。メンブレンを強化化学発光反応溶液(Pierce、Rockford、IL、USA)に1分間浸しました。液体を取り除いた後、サンプルを食品保存フィルムで覆った。開発して修正した後、結果が観察されました。 GAPDHは内部パラメーターであり、タンパク質インプリント画像はImageJ2xソフトウェアによって分析されました。
ONFHのラットモデルの確立
体重300gから350gの90匹のオスのSprague-Dawley(SD)ラット(Shanghai SLAC Laboratory Animal Co.、Ltd.、Shanghai、China)を選択しました。環境は18〜25°Cに設定され、湿度は45〜70%でした。ラットは騒音を避けて別々のケージで飼育された。適応給餌の1週間後、追跡実験が行われました。
モデルは外傷性ONFHによって確立されました。モデリングの方法は次のとおりです。SDラットを腹腔で麻酔した後、脱毛と皮膚の準備を行いました。大腿骨頭を露出させた後、ラットの皮膚を切断し、組織を分離し、丸い靭帯を切断した。ラットの大腿骨頸部の骨膜を削り取り、大腿骨頭周辺の血液供給を破壊した。関節包、筋肉、皮膚を層ごとに縫合し、再度滅菌しました。
動物のグループ化
70匹の成功したモデル化されたSDラットを7つのグループに分け、各グループに10匹のラットを入れました。アゴミール-NCグループ(0.5 mLのmiR-410アゴニストNC(200 nM、広州リボバイオ株式会社、広東省、中国)は、ONFHの確立に成功してから1週間後に股関節の周りに注射されました)、アゴミール-miR-410グループ(0.5 mLのmiR-410アゴニスト(200 nM、広州リボバイオ株式会社、広東省、中国)は、ONFHの確立が成功してから1週間後に股関節の周りに注入されました)、siRNA-NCグループ(0.4mLの沈黙WntのNC -11ベクトル(1×10 9 を含む プラーク形成単位(PFU)(Shanghai GenePharma Co. Ltd.、Shanghai、China)は、ONFHの確立に成功してから1週間後に股関節の周りに注入されました)、Wnt-11-siRNAグループ(0.4 mLのサイレンシングされたWnt-11ベクター(含有1×10 9 PFU)(Shanghai GenePharma Co. Ltd.、Shanghai、China)は、ONFHの確立に成功してから1週間後に股関節の周りに注射されました)、agomir-miR-410 +過剰発現(OE)-NCグループ(0.5mLのmiR-410アゴニストおよび0.4mLのアップレギュレーションされたWnt-11ベクター(1×10 9 を含む)のNC PFU)(Shanghai GenePharma Co. Ltd.、Shanghai、China)は、ONFHの確立に成功してから1週間後に股関節の周りに注射され、agomir-miR-410 + OE-Wnt-11グループ(0.5mLのmiR-410アゴニスト)および0.4mLのアップレギュレーションされたWnt-11ベクター(1×10 9 を含む) PFU)(Shanghai GenePharma Co. Ltd.、Shanghai、China)は、ONFHの確立に成功してから1週間後に股関節の周りに注入されました。一方、正常群(生理食塩水のみを腹腔内に注射した、10匹のラット)を対照として設定した。 ONFHの4週間後、マイクロCT、骨計測分析、X線観察、骨密度測定、および血清カルシウムとリンのレベルの測定が行われました。
マイクロCTテストとオステオメトリクス分析
ラットの右大腿骨頭をマイクロCTマシン(General Electric(GE)Company、米国マサチューセッツ州)に配置し、ランダムに装備された標準ファントムもスキャンしました。スキャンパラメータは次のとおりです。分解能27μm×27μm×27μm、スキャン電流450 mA、スキャン電圧80 kV、シングルスキャン時間88min。ラットの大腿骨頭の微細構造が詳細に観察されました。仕様に照らしてキャリブレーションした後、再構成のためにラットの異なるグループから3つの直方体関心領域(ROI)をランダムに選択しました(0.5×0.5×0.5 cm 3 )。画像処理はマシンソフトウェアGEMicroviewを使用して実行され、ROIは骨計測によって再構築および分析されました。骨塩密度(BMD)と骨体積分率(BV / TV)のパラメーターが選択されました。
X線観察、BMD測定、および血清カルシウムとリンのレベル測定
手術の4週間後、各グループのラットに10%抱水クロラールを腹部から注射しました。麻酔後、ラットを仰臥位にし、四肢を固定し、X線写真で調べた。大腿骨頭の形状と密度の変化が観察されました。左大腿骨背骨、大腿骨頭、および脊椎の骨密度は、二重エネルギーX線骨密度測定器によってテストされました。手術後4週間で、各グループのラットを採血前に12時間絶食させた。血液サンプル(5 mL)を朝に心臓から採取し、真空洗浄した凝固チューブに入れました。血清は、3000 r / minの15分間の遠心分離によって分離されました。血清カルシウムとリンのレベルは、自動生化学分析装置によって測定されました。
電子顕微鏡観察
ラットの骨組織塊を3.5%グルタルアルデヒドで固定し、5%塩酸溶液と1%オスミン酸で脱灰した後、勾配アセトンで脱水し、酢酸ウランとクエン酸で二重染色し、最後にEpon-61を包埋しました。半薄切片を作製した後、透過型電子顕微鏡で超薄切片を観察した。
TdTを介したdUTP-ビオチンニックエンドラベリング(TUNEL)アッセイ
パラフィン包埋切片は、従来の脱ロウおよび脱水で前処理された。切片をペプシン(0.25〜0.5%塩酸溶液)で25分間ハッチングした後、50μLのTUNEL反応混合溶液を滴下し、ウェットボックスで60分間ハッチングし、50μLの薬剤-ペルオキシダーゼを滴下してハッチングしました。 30分間ウェットボックス。切片が着色され、顕微鏡下で観察されたかどうかにかかわらず、切片はDAB試薬で現像された。切片に水を加えて発育を停止し、ヘマトキシリンで2分間染色した。次に、切片を95%エタノールI–II、無水エタノールI–IIにそれぞれ3–5分、キシレンI–IIに3–5分浸し、中性ガムで密封しました。結果は光学顕微鏡で分析されました。
酵素結合免疫吸着測定法(ELISA)
ラットに麻酔をかけた後、大腿動脈の血液を採取し、1時間の休止後に遠心分離によって血清サンプルを収集した。 ELISAキット(上海酵素結合バイオテクノロジー株式会社、上海、中国)の仕様に基づいて、7つの標準ウェルにさまざまな濃度標準(100μL)を順番に追加し、ブランクウェルに100μLを追加しました。標準希釈液、および残りのウェルにテストする100μLのサンプルを追加しました。酵素プレートをフィルムでコーティングし、1時間置いた後、液体を廃棄した。溶液(100μL)をすべてのウェルに添加し、酵素プレートをフィルムで1時間覆った。洗浄後、100μLのB溶液をすべてのウェルに加え、酵素プレートをフィルムで30分間覆った。次に、テトラメチルベンジジン基質溶液(90μL)を加え、酵素プレートをフィルムで10〜20分間覆い、50μLの終結溶液をすべてのウェルに加えました。各ウェルの光学密度(OD)値をマイクロプレートリーダーで測定し、サンプルの実際の濃度を列挙しました。
デュアルルシフェラーゼレポーター遺伝子アッセイ
miR-410とWnt-113 '非翻訳領域(UTR)の結合部位と標的関係は、バイオインフォマティクスソフトウェアhttp://www.targetscan.orgを使用して予測されました。 miR-410結合部位を含むWnt-113'UTRプロモーター領域配列を構成し、pMIR-REPORT TM に挿入しました。 Wnt-11 3'UTR野生型プラスミド(Wnt-11-WT)を処方するためのルシフェラーゼベクタープラスミド(Ambion、Company、Austin、TX、USA)。そして、Wnt-11 3'UTR変異プラスミド(Wnt-11-MUT)は、プラスミドと変異結合部位に基づいて構築されました。購入したプラスミド抽出キット(Promega、マディソン、ウィスコンシン、米国)の手順に従い、対数細胞を96ウェルプレートに播種しました。細胞のコンフルエンスが約70%になったとき、Lipofectamine2000をトランスフェクションに採用しました。 Wnt-11-WTおよびWnt-11-MUTは、それぞれミミックNCおよびmiR-410ミミック(Shanghai GenePharma Co. Ltd.、Shanghai、China)と混合され、293T細胞にコトランスフェクトされました。トランスフェクト48時間後に細胞を集めて溶解し、ルシフェラーゼ検出キット(BioVision、サンフランシスコ、カリフォルニア州、米国)およびGlomax 20/20ルミノメーター(Promega、マディソン、ウィスコンシン州、米国)によってルシフェラーゼ活性をテストしました。
統計分析
すべてのデータは、SPSS 21.0ソフトウェア(IBM Corp. Armonk、NY、USA)によって処理されました。測定データは平均±標準偏差で伝えられました。一元配置分散分析(ANOVA)は、複数グループの比較と t のために実施されました。 2つのグループの比較とフィッシャーの最小有意差 t をテストします テスト(LSD-t)はANOVAの後に利用されました。 P 値<0.05は、統計的に有意な差を示していました。
結果
臨床サンプルのONFH組織の病理学的変化
HE染色は、大腿骨頸部骨折群(対照群)とONFH群の大腿骨頭の病理学的変化を観察するために採用されました。結果は、対照群では、骨梁密度がよく分布しており、構造が完全であったことを報告しました。 ONFHグループでは、小脳に空の骨小脳が多く、骨細胞が減少し、骨梁が継続的に変化していました。さらに、骨の小脳の周りには他にも多くの組織過形成がありました(図1a)。
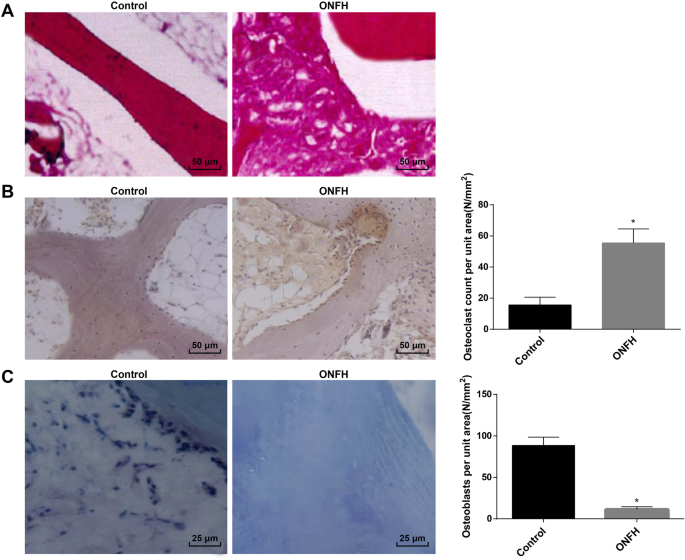
臨床サンプルのONFH組織の病理学的変化。 a 各グループ(200×)でのHE染色の結果。 b TRAP染色の結果と各グループ(200×)のTRAP陽性細胞の数。 c ALP染色およびALP陽性細胞の結果は各グループ(400×)でカウントされます。 * P <0.05対対照群
TRAP染色により、TRAPは主に破骨細胞に見られ、アマランサスの成熟破骨細胞を特定するためによく使用されることが明らかになりました。対照群と比較して、ONFH群のTRAP陽性破骨細胞の数は増加し、破骨細胞の形態は多様であり、細胞は大きく、多核の外観を示した。破骨細胞に隣接する骨小脳に明らかな骨欠損が観察され、骨吸収が見られました(図1b)。
ALP染色の結果は、ALPが骨芽細胞の分化と成熟のマーカー酵素であり、骨基質の成熟石灰化の調節に関与していることを示しました。したがって、ALP発現は骨芽細胞を同定するために一般的に使用されました。成熟した骨芽細胞の細胞質に茶色またはコーヒーの粒子が見られた。対照群と比較して、ALP陽性骨芽細胞の数はONFH群で減少しました(図1c)。
MiR-410、ALP、BGLAP、およびCollα1が分解され、Wnt-11、ACP5、CTSK、およびMMP9が臨床サンプルのONFH組織で強化されました
>ウエスタンブロット分析およびRT-qPCRは、ONFHグループのmiR-410発現が対照グループのそれと比較して減少した一方で、Wnt-11発現が上昇したことを示しました。骨芽細胞関連因子ALP、BGLAP、およびCollα1の発現が低下しました。破骨細胞関連因子ACP5、CTSK、およびMMP9の発現が上昇しました(すべて P <0.05)(図2a–c)。
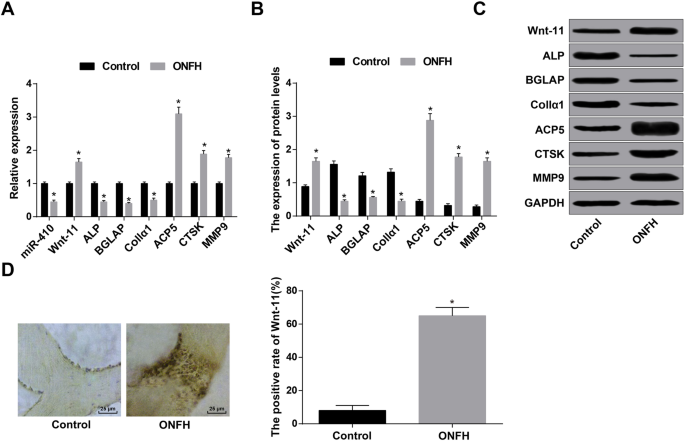
臨床サンプルのONFH組織では、MiR-410、ALP、BGLAP、およびCollα1が減少し、Wnt-11、ACP5、CTSK、およびMMP9が増加しました。 a RT-qPCRによるmiR-410、Wnt-11、骨芽細胞および破骨細胞関連遺伝子の検出。 b ウエスタンブロット分析によって検出されたWnt-11および骨芽細胞および破骨細胞関連タンパク質。 c Wnt-11および骨芽細胞および破骨細胞関連タンパク質のタンパク質バンド。 d Wnt11発現(400×)および免疫組織化学的染色によって検出されたWnt11タンパク質陽性率の比較。 * P <0.05対対照群
Wnt-11の発現は免疫組織化学によってテストされ、結果はWnt-11タンパク質が主に細胞質で発現され、陽性の発現は基本的に茶色がかった黄色または茶色であることを示しました。対照群とは対照的に、ONFH群におけるWnt-11タンパク質発現の陽性率は上昇しました( P <0.05)(図2d)。
ダウンレギュレーションされたmiR-410とダウンレギュレーションされたWnt-11はラットのBMDとBV / TVを上昇させます
マイクロCTは、正常群では、大腿骨頭の外観は丸く、首の厚さは均一であり、骨梁の構造は連続的で均一に分布していることを示唆しました。 ONFH群、agomir-NC群、siRNA-NC群、agomir-miR-410 + OE-Wnt-11群のラットの大腿骨頭は徐々に崩壊し、骨吸収領域は徐々に現れ、大腿骨の首は薄くなった。 、そして骨梁が壊れ、連続性が破壊されました。アゴミール-miR-410グループ、Wnt-11-siRNAグループ、およびアゴミール-miR-410 + OE-NCグループでは、ラットの外観は無傷のままであり、崩壊は起こらず、明らかな骨吸収領域は現れず、骨梁が見られた。正常に配置され、構造が完成しました(図3a)。
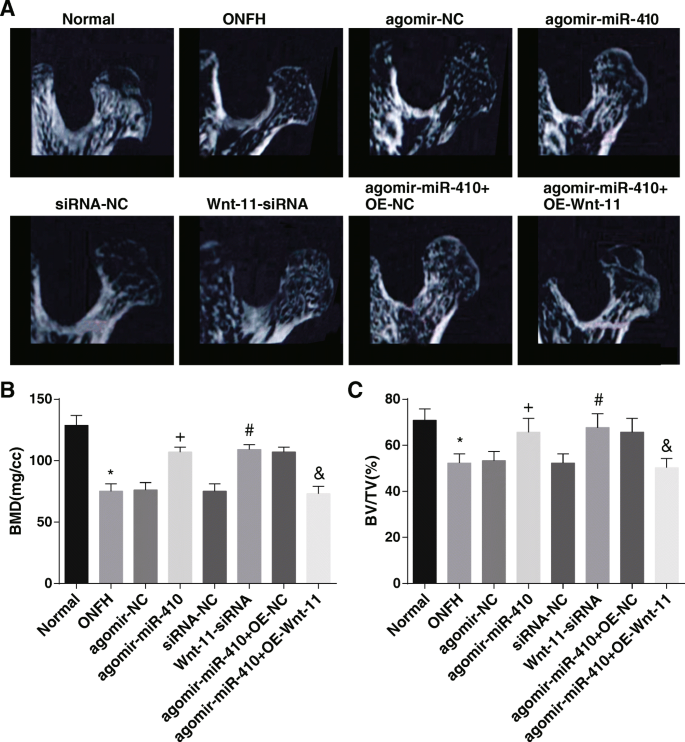
高度に発現されたmiR-410と不十分に発現されたWnt-11は、ラットのBMDとBV / TVを増加させます。 a 各グループのラットのマイクロCT結果。 b BMD分析の結果。 c BV / TV分析の結果。 * P <0.05対通常のグループ。 + P <0.05対アゴミール-NCグループ。 # P <0.05 vs.siRNA-NCグループ。 & P <0.05vs。アゴミール-miR-410 + OE-NCグループ
骨計測の結果は、正常なグループとは対照的に、ONFHグループのBMDとBV / TVが落ち込んでいることを報告しました(両方 P <0.05)。アゴミール-NCグループとsiRNA-NCグループに関連して、アゴミール-miR-410グループとWnt-11-siRNAグループ(すべて P )でBMDとBV / TVが上昇しました。 <0.05)。 agomir-miR-410 + OE-NCグループと比較すると、agomir-miR-410 + OE-Wnt-11グループのBMDとBV / TVは削除されました(両方 P <0.05)(図3b、c)。
miR-410の過剰発現とWnt-11の発現不良により、大腿骨シャフト、大腿骨頭、脊柱のBMDレベルが上昇し、血清カルシウムも上昇します。ラットのリンレベルとリンレベル
X線検査では、正常群の大腿骨頭の外観は丸く、大腿骨頭の密度は均一で、構造は完全であることが観察されました。 ONFH群、agomir-NC群、siRNA-NC群、agomir-miR-410 + OE-Wnt11群では、ラットの大腿骨頭の外観が薄くなり、骨吸収領域が現れた。外観は均一で、ラットの大腿骨頭の厚さは、agomir-miR-410グループ、Wnt11-siRNAグループ、およびagomir-miR-410グループ+ OE-NCグループで完全でした(図4a)。 P>
上方制御されたmiR-410と下方制御されたWnt-11は、大腿骨頭、大腿骨頭、脊柱の骨塩密度レベルを上昇させ、ラットの血清カルシウムとリンのレベルも上昇させます。 a 動物のX線観察結果。 b 各グループの大腿骨シャフト、大腿骨頭、および脊椎の骨塩密度の比較。 c 各グループの血清カルシウムおよびリンレベルの比較。 * P <0.05対通常のグループ。
+
P <0.05対アゴミール-NCグループ。
#
P <0.05 vs.siRNA-NCグループ。
&
P <0.05vs。アゴミール-miR-410 + OE-NCグループ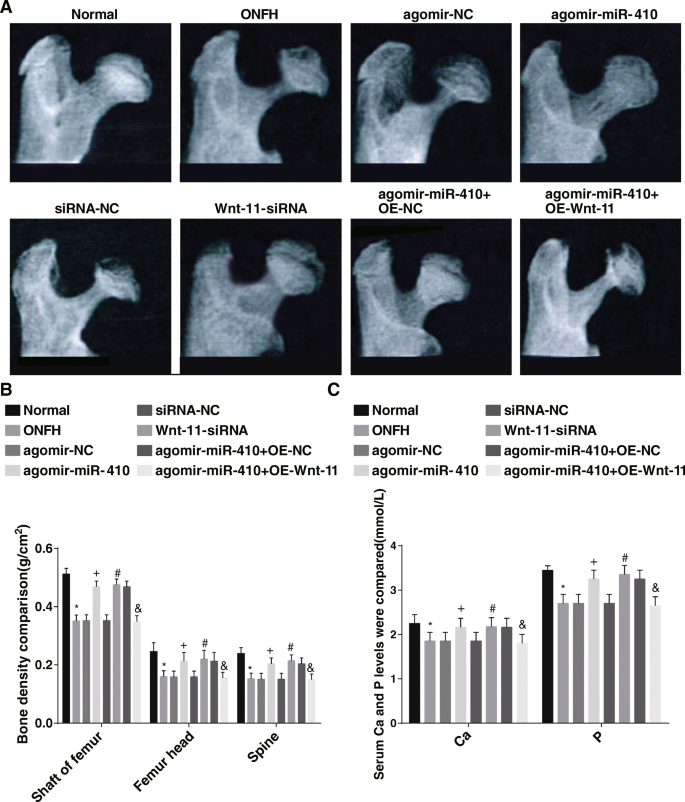
BMDと血清カルシウムおよびリンレベルの測定では、正常グループと比較して、大腿骨頭、大腿骨頭、脊椎のBMD、および血清カルシウムとリンのレベルがONFHグループで低下したことが報告されました(すべて P <0.05)。アゴミール-NCグループおよびsiRNA-NCグループとは対照的に、大腿骨頭、大腿骨頭、および脊椎のBMD、ならびに血清カルシウムおよびリンレベルは、アゴミール-miR-410グループおよびWnt-11-で上昇しました。 siRNAグループ(すべて P <0.05)。アゴミール-miR-410 + OE-NCグループに関連して、大腿骨シャフト、大腿骨頭、脊椎のBMD、およびアゴミール-miR-410 + OE-Wnt-11の血清カルシウムとリンのレベルグループ、軽減されました(すべての P <0.05)(図4b、c)。
Wnt-11のサイレンシングとmiR-410のアップレギュレーションにより、ラット組織の病理学的変化が緩和され、骨細胞のアポトーシスが抑制されます
HE染色の結果は、骨梁がきちんとしていて透明であり、規則的かつしっかりと配置されていることを示した。骨細胞は骨の裂孔を満たし、石灰化したゾーンは正常なグループの軟骨下骨梁によく接続されていました。 ONFHグループ、agomir-NCグループ、siRNA-NCグループ、およびagomir-miR-410 + OE-Wnt-11グループでは、骨梁はまばらでした。薄くなり、壊れさえします。構造が乱れていました。そして、いくつかの断片が現れました。骨小柱の一部の骨細胞は壊死性であり、多数の骨小窩は空であり、骨細胞は充填されておらず、壊死性骨小柱の周りに巻き付けられた壊死骨小柱空間で肉芽組織の明らかな増殖が観察された。アゴミール-miR-410グループ、Wnt-11-siRNAグループ、アゴミール-miR-410 + OE-NCグループでは、ラットの外観は損なわれず、明らかな骨吸収領域は見られず、骨梁が配置されていました。通常、構造は完成しました(図5a)。
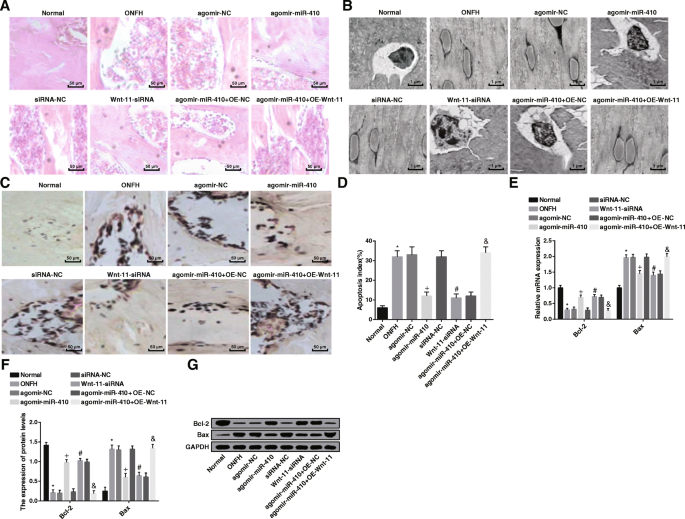
Wnt-11の不十分な発現とmiR-410の過剰発現は、ラット組織の病理学的変化を軽減し、骨細胞のアポトーシスを抑制します。 a 各群(200×)のラットの大腿骨頭の形態学的観察。 b 各群(8000×)のラットの骨細胞の超微細構造観察。 c TUNEL染色の結果(200×)。 d TUNELアッセイによるアポトーシス率の検出。 e RT-qPCRによるBaxおよびBcl-2mRNA発現の検出。 f 、 g ウエスタンブロット分析によるBaxおよびBcl-2タンパク質発現の検出。 * P <0.05対通常のグループ。 + P <0.05対アゴミール-NCグループ。 # P <0.05 vs.siRNA-NCグループ。 & P <0.05vs。アゴミール-miR-410 + OE-NCグループ
電子顕微鏡観察により、正常群の骨細胞の形状は裂孔と一致しており、細胞と裂孔壁との間に小さなギャップがあったことが観察された。細胞小器官は豊富で、小器官の細胞は正常であり、核は卵形であり、核膜は無傷であり、クロマチンは凝集せず、細胞質の偽足は末梢骨の小管に伸びて、隣接する骨細胞。骨芽細胞は骨梁の表面にあり、サンプルは長く、細胞小器官が多かった。 ONFHグループ、agomir-NCグループ、siRNA-NCグループ、およびagomir-miR-410 + OE-Wnt-11グループでは、骨芽細胞の細胞質、カプセルと裂孔の間のギャップに多数の脂質沈着物が見られました。さらに拡大し、核膜と細胞質の間に浮腫の明るいバンドが現れ、核膜が圧迫されて現れ、核膜は無傷で、細胞質のミトコンドリアは腫れ、小胞体とゴア装置は消えた。アゴミール-miR-410グループ、Wnt-11-siRNAグループ、アゴミール-miR-410 + OE-NCグループでは、ラット骨芽細胞の形態は正常であり、少数の細胞の核にクロマチン凝集が見られました。細胞質には明らかな脂肪滴は見られず、核膜は無傷でした(図5b)。
TUNEL染色の結果は、正常群と比較して、ONFH群の骨細胞のアポトーシス率が上昇したことを示しました( P <0.05)。アゴミール-NCグループおよびsiRNA-NCグループとは対照的に、骨細胞のアポトーシス率は、アゴミール-miR-410グループおよびWnt-11-siRNAグループ(両方 P )で減少しました。 <0.05)。アゴミール-miR-410 + OE-NCグループに関連して、アゴミール-miR-410 + OE-Wnt-11グループ( P )では骨細胞のアポトーシス率が上昇しました。 <0.05)(図5c、d)。
ウエスタンブロット分析とRT-qPCRは、正常群と比較してONFH群でBcl-2発現が減少し、Bax発現が上昇したことを報告しました(両方 P <0.05)。アゴミール-NCグループとsiRNA-NCグループと比較すると、アゴミール-miR-410グループとWnt-11-siRNAグループ(すべて P )ではBcl-2発現が上昇し、Bax発現が下降しました。 <0.05)。アゴミール-miR-410 + OE-NCグループとは対照的に、アゴミール-miR-410 + OE-Wnt-11グループ(両方 P )では、Bcl-2発現が低下し、Bax発現が追加されました。 <0.05)(図5e–g)。
miR-410の過剰発現とWnt-11の低発現は、骨芽細胞の数を増やし、破骨細胞の数を減らします
TRAP染色により、正常群では、TRAP染色陽性の破骨細胞の形態が異なり、それらのほとんどがシャトルまたは紡錘状であり、いくつかの大きな破骨細胞が多角形であり、それらのほとんどが小脳の周りに分布していたことが明らかになった。 ONFH群、agomir-NC群、siRNA-NC群、agomir-miR-410 + OE-Wnt-11群では、TRAP陽性細胞が上昇し、細胞の形態は大きな多角形と多核細胞で多様であり、骨吸収破骨細胞に隣接する骨梁で発見され、典型的な吸収性裂孔が形成された。アゴミール-miR-410グループ、Wnt-11-siRNAグループ、アゴミール-miR-410 + OE-NCグループでは、TRAP染色陽性の細胞数が減少し、細胞は長い帯状になり、形態が変化しました。より規則的であり、多角形の多核破骨細胞はまれであり、隣接する骨梁の骨構造は比較的完全でした(図6a)。
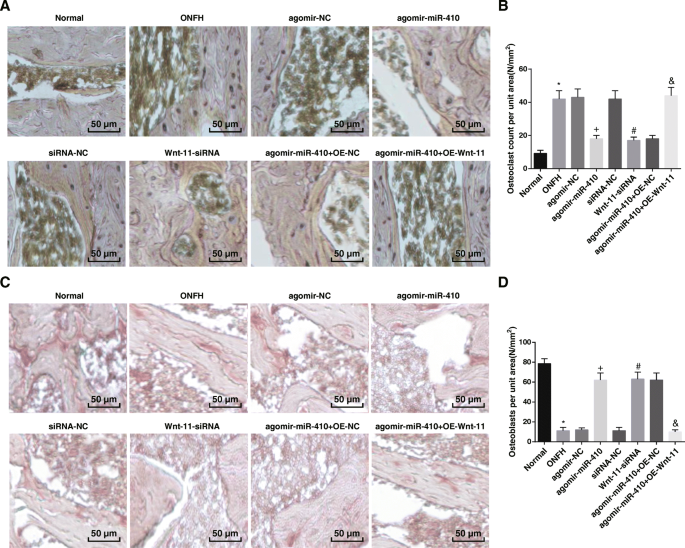
miR-410の過剰発現とWnt-11の低発現は、骨芽細胞の数を増やし、破骨細胞の数を減らします。 a ラット(200×)の破骨細胞におけるTRAP染色の結果。 b TRAP染色における陽性細胞の数。 c 各群(200×)のラットの骨芽細胞におけるALP染色の結果。 d ALP染色によって決定された陽性細胞の数。 * P <0.05対通常のグループ。 + P <0.05対アゴミール-NCグループ。 # P <0.05 vs.siRNA-NCグループ。 & P <0.05vs。アゴミール-miR-410 + OE-NCグループ
ALP染色は、正常群において、ALP染色に対して陽性であった骨芽細胞において、形態が小さく、丸いことを示した。細胞は凝集して分布し、そのほとんどは骨髄腔の骨梁空間といくつかの骨梁の表面に位置していた。 ONFH群、agomir-NC群、siRNA-NC群、agomir-miR-410 + OE-Wnt-11群では、ALP染色陽性の骨芽細胞の数が骨髄腔の小柱空間に分散して分布していた。アゴミール-miR-410グループ、Wnt-11-siRNAグループ、アゴミール-miR-410 + OE-NCグループでは、骨芽細胞陽性細胞の数が増加し、骨髄腔の小柱空間に分布していた。骨梁の一部の表面(図6c)。
正常群と比較して、ONFH群( P )では単位面積あたりの骨芽細胞数が抑制され、破骨細胞数が増加した。 <0.05)。アゴミール-NC群とsiRNA-NC群では、アゴミール-miR-410群とWnt-11-siRNA群で単位面積あたりの骨芽細胞数が増加し、破骨細胞数が減少した(すべて P <0.05)。アゴミール-miR-410 + OE-NCグループとは対照的に、アゴミール-miR-410 + OE-Wnt-11グループでは、単位面積あたりの骨芽細胞数が減少し、破骨細胞数が増加しました(両方 P <0.05)(図6b、d)。
高発現miR-410および低発現Wnt-11は、オステオカルシン(OCN)、ALP、BGLAP、およびCollα1、ならびにAbateC-の発現を上昇させますONFHラットにおけるI型コラーゲン(NTX-1、CTX-1)、ACP5、CTSK、およびMMP9発現のN末端テロペプチド
ELISAの結果は、正常群と比較して、破骨細胞機能指数NTX-1およびCTX-1のレベルがONFH群で上昇する一方で、骨形成機能指数ALPおよびOCNのレベルが低下することを明らかにしました(すべて P <0.05)。アゴミール-NCグループとsiRNA-NCグループとは対照的に、ALPとOCNは上昇し、NTX-1とCTX-1はアゴミール-miR-410グループとWnt-11-siRNAグループで下降しました(すべて P <0.05)。 agomir-miR-410 + OE-NCグループと比較すると、ALPとOCNはドロップし、NTX-1とCTX-1はagomir-miR-410 + OE-Wnt-11グループに追加されました(すべて P <0.05)(図7a)。
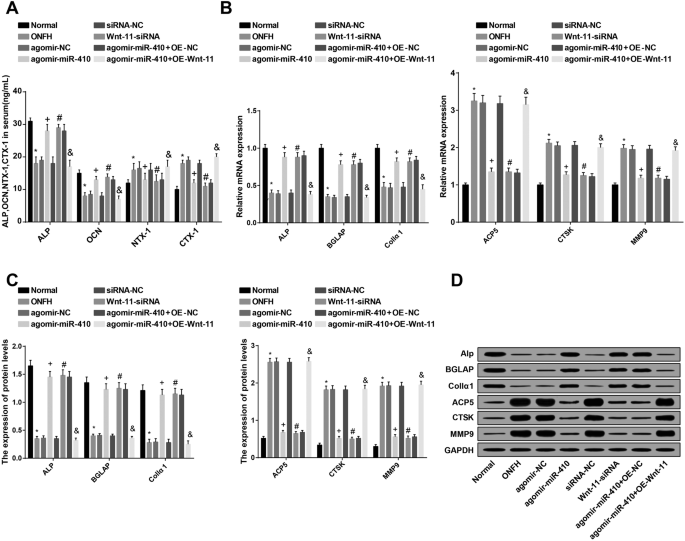
高発現miR-410および低発現Wnt-11は、OCN、ALP、BGLAP、およびCollα1の発現を上昇させ、ONFHラットにおけるNTX-1、CTX-1、ACP5、CTSK、およびMMP9の発現を抑制します。 a ELISAによる各群のラットの血清中の骨形成および破骨細胞機能指数の検出。 b RT-qPCRによる骨芽細胞および破骨細胞関連遺伝子の検出。 c 、 d ウエスタンブロット分析による骨芽細胞および破骨細胞関連遺伝子のタンパク質発現の検出。 * P <0.05対通常のグループ。 + P <0.05対アゴミール-NCグループ。 # P <0.05 vs.siRNA-NCグループ。 & P <0.05vs。アゴミール-miR-410 + OE-NCグループ
ウエスタンブロット分析およびRT-qPCRの結果は、正常群と比較して、骨芽細胞関連因子ALP、BGLAP、およびCollα1の発現が低下し、破骨細胞関連因子ACP5、CTSK、およびMMP9の発現が上昇したことを明らかにしました(すべて P <0.05)。アゴミール-NCグループおよびsiRNA-NCグループとは対照的に、ALP、BGLAP、およびCollα1の発現は上昇し、ACP5、CTSK、およびMMP9の発現はアゴミール-miR-410グループおよびWnt-11-siRNAで減少しました。グループ(すべて P <0.05)。アゴミール-miR-410 + OE-NC群と比較すると、アゴミール-miR-410 + OE-Wnt-11群ではALP、BGLAP、Collα1の発現が抑制され、ACP5、CTSK、MMP9の発現が上昇した(すべての P <0.05)(図7b–d)。
Wnt-11の上昇とmiR-410の低下はラットのONFH組織に見られ、Wnt-11はmiR-410の標的遺伝子です
>miR-410とWnt-11遺伝子のターゲティング関係は、オンライン分析ソフトウェアによって分析されました。 Wnt-11遺伝子配列とmiR-410配列の間に特異的結合領域が存在することが示され、Wnt-11がmiR-410の標的遺伝子であることが示唆されました(図8a)。ルシフェラーゼ活性アッセイを利用して、この関係を検証しました(図8b)。結果は、NCグループと比較して、miR-410ミミックグループ( P )でルシフェラーゼ活性が低下したことを示しました。 <0.05)、しかしMUT 3'UTR( P )のルシフェラーゼ活性に有意差はありませんでした> 0.05)、miR-410がWnt-11に特異的に結合できることを示しています。
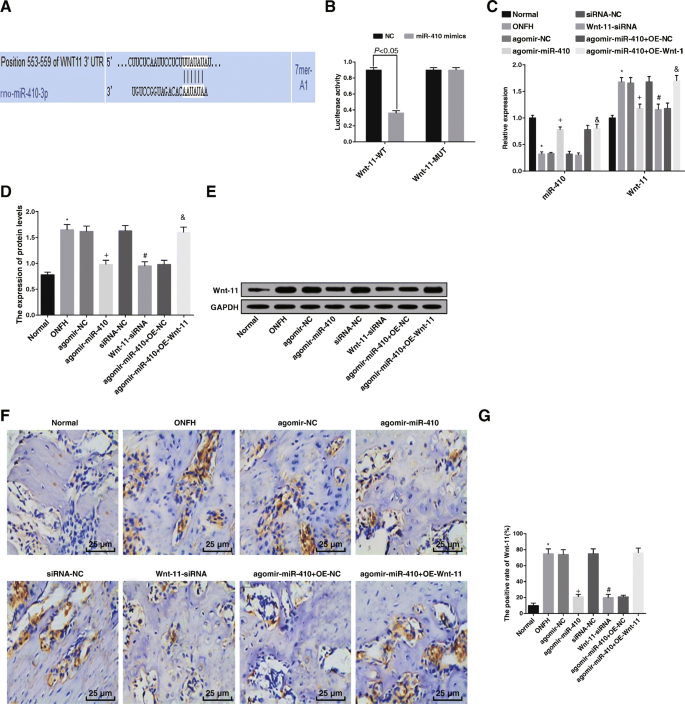
Wnt-11の増加とmiR-410の減少は、ONFH組織に見られ、Wnt-11はmiR-410の標的遺伝子です。 a Wnt-113'UTR上のmiR-410の結合部位の予測。 b miR-410とWnt-11の関係を検証するためのルシフェラーゼ活性検出。 c RT-qPCRによるmiR-410およびWnt-11発現の検出。 d 、 e ウエスタンブロット分析によって決定されたWnt-11タンパク質発現。 f 免疫組織化学(200×)によって検出された各グループのラットの大腿骨頭におけるWnt-11の発現。 g 各グループにおけるWnt-11タンパク質発現の陽性率の比較。 * P <0.05対通常のグループ。 + P <0.05対アゴミール-NCグループ。 # P <0.05 vs.siRNA-NCグループ。 & P <0.05vs。アゴミール-miR-410 + OE-NCグループ
ウエスタンブロット分析とRT-qPCRの結果は、正常群と比較して、ONFH群( P の両方)でmiR-410の発現が低下し、Wnt-11の発現が上昇したことを明らかにしました。 <0.05)。アゴミール-NCグループと比較すると、アゴミール-miR-410グループではmiR-410が上昇し、Wnt-11が低下しました(両方 P <0.05)。 siRNA-NCグループとは対照的に、Wnt-11はWnt-11-siRNAグループで減少しました( P <0.05)。 Wnt-11は、agomir-miR-410 + OE-NCグループ( P )と比較して、agomir-miR-410 + OE-Wnt-11グループで強化されました。 <0.05)(図8c–e)。
> Wnt-11の発現は、免疫組織化学によって確認されました。 Wnt-11タンパク質は主に細胞質で発現し、陽性発現は主に褐色がかった黄色または褐色でした。正常群と比較して、ONFH群のWnt-11タンパク質発現の陽性率が上昇しました( P <0.05)。アゴミール-NCグループおよびsiRNA-NCグループと比較すると、Wnt-11タンパク質発現の陽性率は、アゴミール-miR-410グループおよびWnt-11-siRNAグループ(両方 P > <0.05)。アゴミール-miR-410 + OE-NCグループに関連して、アゴミール-miR-410 + OE-Wnt-11グループ( P )ではWnt-11タンパク質発現の陽性率が向上しました。 <0.05)(図8f、g)。
ディスカッション
骨破壊疾患の一種であるONFHは、血液供給の失敗、凝固、線維素溶解系の障害によって引き起こされ、最終的に大腿骨頭が崩壊します[19]。以前の研究では、ONFHのマウスでホルモンによって誘発された骨髄間葉系幹細胞でのmiRNA発現について議論されています[20]。また、最近の研究は、ステロイド誘発性ONFHに関連する骨形成における骨髄由来間葉系幹細胞のmiRNAの発現プロファイルの証拠を提供しました[21]。さらに、Li-nHA / GM / rhEPOステントはWnt経路とHIF-1 / VEGF経路の両方を上昇させ、糖質コルチコイドによって誘発されるONFHの修復に有益な骨形成と血管新生を促進できることが明らかになりました[22]。これらの事実に基づいて、この研究は、Wnt-11を標的とすることにより、ONFHの予防におけるmiR-410の効果を調査することを目的としました。
私たちの研究は、miR-410、ALP、BGLAP、およびCollα1がONFH組織で分解され、Wnt-11、ACP5、CTSK、およびMMP9が増強されるという概念に関連する実質的な証拠を提供しました。最近の研究では、骨肉腫におけるmiR-410の発現が低下することが促進されており、抗腫瘍効果が示されています[23]。別の研究では、miR-410の発現が乳がんのヒトエストロゲン受容体陽性組織で減少したことが示されています[24]。分泌因子Wnt-11の発現は、結腸直腸癌を含むいくつかのタイプの癌で上昇し、癌細胞の移動と浸潤を促進すると報告されています[25]。同様に、以前の研究では、ホルモン非依存性の前立腺癌でWnt-11の発現が高まることが証明されています[26]。 ALPは、疾患の状態診断および臨床的予後のための有用な指標です[27]。 OCN(骨γ-カルボキシグルタミン酸タンパク質; BGLAP)は、骨基質の鉱化作用に関連する広く保存されている分子です[28]。 ACP5は破骨細胞の分化と骨吸収に必要であり、接着キナーゼのリン酸化を調節することで細胞の動きを促進します[29]。 CTSKは、オステオポンチン、I型コラーゲンおよびその他の骨基質タンパク質を分解する重要なプロテアーゼです[30]。 MMPは、細胞外マトリックスと基底膜のほとんどの成分を分解および修飾し、癌の浸潤と転移に計り知れない影響を与える可能性があります[31]。また、我々の研究は、Wnt-11がmiR-410の標的遺伝子であることを明らかにしました。同様に、張等。 miR-410がWnt3a3'-UTRの推定結合部位を標的にして、Wntシグナル伝達経路を調節することを発見しました[32]。
さらに、miR-410のアップレギュレーションとWnt-11のダウンレギュレーションにより、ONFHラットのBMDとBV / TVが増加することが明らかになりました。非外傷性ONFH患者における脊椎BMDの減少と尿中DPD / Cr比の増加が以前に示唆された[33]。別の研究では、ONFHグループの大腿骨頭と腰椎のBMDが対照グループのBMDと比較して低下していることが確認されています[34]。さらに、画像解析により、ムスコンは壊死した大腿骨頭のBMDおよびBV / TV比を回復できることが明らかになり、組織学的検査により、アルコール誘発性ONFHに対するムスコンの保護効果がさらに確認されました[35]。 miR-410を高発現し、Wnt-11を低発現すると骨細胞のアポトーシスが抑制されるという結果が得られました。ある研究では、miR-410をサイレンシングすると、細胞増殖が誘導され、酸化低密度リポタンパク質によって誘導されるヒト臍帯静脈内皮細胞のアポトーシスが減少することが示されています[36]。下降したmiR-410の発現と上昇したSOCS3の発現は、抗アポトーシス因子Bcl-2の発現を低下させ、細胞のアポトーシスを促進する可能性があると報告されています[37]。さらに、アンドロゲン欠損LNCaP細胞では、Wnt-11のダウンレギュレーションが神経内分泌様の分化を防ぎ、前立腺癌細胞のアポトーシスを引き起こす可能性があります[26]。この研究はまた、Wnt-11のダウンレギュレーションとmiR-410のアップレギュレーションが、ALP、BGLAP、およびCollα1の発現を増強し、ACP5、CTSK、およびMMP9の発現を減少させることを示しました。ある研究では、子宮頸がん細胞で高められたWnt-11発現は、Wnt / Jnk経路を活性化することにより、JNK-1の活性化とリン酸化を引き起こし、腫瘍細胞の増殖と遊走/浸潤を促進する可能性があることが示されています[15]。結腸直腸癌細胞株におけるWnt-11遺伝子サイレンシングは細胞の浸潤能を低下させることが示唆されています[25]。さらに、miR-410の過剰発現は、TRIM44を直接標的とすることにより、骨肉腫細胞の浸潤、遊走、増殖、および上皮間葉転換を抑制することができます[23]。さらに、以前の研究では、アップレギュレーションされたmiR-410は、アポトーシスを誘導することにより、異種移植マウスモデルにおける胆管癌の増殖を抑制したと述べています[38]。
結論
要約すると、我々の調査は、miR-410が低発現し、Wnt-11がONFHで高発現し、miR-410をアップレギュレーションまたはWnt-11をダウンレギュレーションすると、骨芽細胞が増加し、破骨細胞が減少してONFHが軽減されることを明らかにしました。ただし、ONFHの治療におけるmiR-410およびWnt-11の有効性を検出するために、臨床研究がさらに実施される可能性があります。
データと資料の可用性
該当なし
変更履歴
略語
- ALP:
-
アルカリホスファターゼ
- ANOVA:
-
分散分析
- BMD:
-
骨密度
- EDTA:
-
エチレンジアミン四酢酸
- ELISA:
-
酵素免疫測定法
- GAPDH:
-
グリセルアルデヒドリン酸デヒドロゲナーゼ
- GE:
-
ゼネラルエレクトリック
- HE:
-
ヘマトキシリン-エオシン
- LSD-t:
-
最小有意差 t テスト
- miR-410:
-
MicroRNA-410
- miRNAs:
-
マイクロRNA
- NC:
-
ネガティブコントロール
- OD:
-
光学密度
- ONFH:
-
大腿骨頭の骨壊死
- PBS:
-
リン酸緩衝生理食塩水
- ROI:
-
関心のある地域
- RT-qPCR:
-
逆転写定量的ポリメラーゼ連鎖反応
- SD:
-
Sprague-Dawley
- SDS:
-
ドデシル硫酸ナトリウム
- THR:
-
人工股関節全置換術
- TRAP:
-
酒石酸耐性酸性ホスファターゼ
- TUNEL:
-
TdTを介したdUTP-ビオチンニックエンドラベリング
ナノマテリアル
- スタックカップカーボンナノファイバーの原子および電子構造を明らかにする
- ボロフェンの安定性とSTM画像に関する第一原理研究
- 微結晶およびナノセルロースの構造と誘電特性に及ぼす水の影響
- 界面層の設計によるZnO膜の表面形態と特性の調整
- メタマテリアルにおける表面プラズモンポラリトンと磁気双極子共鳴の結合効果
- Ag n V(n =1–12)クラスターの構造的、電子的、および磁気的特性の調査
- ポリオール媒介プロセスによるZnOナノクリップの製造と特性評価
- 小胞および細胞の周波数変調波誘電泳動:クロスオーバー周波数での周期的Uターン
- 低エネルギー照射に対するSi、Ge、およびSi / Ge超格子の放射応答の理論的シミュレーション
- チタニアナノチューブの熱伝導率に及ぼす形態と結晶構造の影響
- LSPカップリングの波長と強度に対する基板の影響