


金ナノロッドコア/銀シェルナノ構造によって誘発される遺伝子毒性におけるROSと細胞周期停止の役割
要約
銀ナノ粒子(AgNPs)と銀イオンによって肝臓に誘発される遺伝子毒性を理解するために、この研究では、操作された金ナノロッドコア/銀シェルナノ構造(Au @ Ag NR)とヒト化肝細胞HepaRG細胞を使用しました。 0.4〜20 µg mL -1 によって誘発されるDNAおよび染色体損傷における酸化ストレスおよび細胞周期停止の関与 Au @ Ag NRは、コメットアッセイ、γ-H2AXアッセイ、および小核試験によって調査されました。さらに、Au @ AgNRの分布を分析しました。私たちの結果は、Ag + の両方が また、Au @ Ag NRはHepaRG細胞のDNA切断と染色体損傷(染色体異常誘発性)を引き起こし、核に保持されたAu @ AgNRはさらにAg + を放出する可能性があります。 、主に細胞周期の停止とROSの形成によって引き起こされる損傷を悪化させます。結果は、細胞内蓄積、Ag + 間の相関関係を明らかにします イオン放出とAgNPの潜在的な遺伝子毒性。
はじめに
サイズが1〜100 nmの銀ナノ粒子(AgNP)は、病原体に侵入し、それらの代謝酵素の内部スルフヒドリル基を不活性化することにより、広範囲の抗菌特性を示すことができます[1]。彼らは、 Escherichia Coli で強力な静菌作用と殺菌効果を示しました。 、 Neisseria gonorrhoeae およびクラミジアトラコマチス 、および医療用コーティング、家庭用製品[2]、および創傷被覆材[3]として広く使用されています。説得力のある証拠は、ナノ粒子が核に入り、DNAの合成および転写プロセスを妨害することができることを示しています[4]。以前の研究では、5 mg / kg AgNPの単回静脈内投与により、Sprague-Dawleyラットの骨髄細胞に顕著な染色体破壊が生じる可能性があることを報告しました[5]。 10 mg / kg以上のAgNPを1回腹腔内注射すると、DNAと染色体の両方の損傷が誘発されました[6]。フラワー他[7]は、50および100μgmL -1 の用量のAgNPを提案しました。 投与後5分以内にDNA損傷を引き起こす可能性があり、急速に放出される銀(Ag)の遺伝子毒性を浮き彫りにします。過度の曝露のリスクを考慮して、ナノ遺伝子毒性学またはDNA損傷と、人工ナノ材料の発がん性の可能性の調査が大きな注目を集めています[8]。
AgNPによって誘発される遺伝的損傷の主なメカニズムは、活性酸素種の過剰産生、炎症、および細胞周期障害であると考えられています[9、10]。以前の研究で示唆されているように、AgNPは酸化的損傷を介してDNAと直接相互作用し[11]、DNAレベルで間期に干渉し、染色体レベルで有糸分裂に干渉するか、核タンパク質および有糸分裂紡錘体装置と相互作用して細胞周期チェックポイントを妨害する可能性があります[11]。 12]。ただし、AgNPによって誘発される遺伝子毒性が、部分的にナノ粒子に起因するのか[13、14]、完全に放出されたAg + に起因するのか イオンはまだ不明です[15、16]。
組織内の銀の放出が不安定で中断されないため、AgNPの遺伝子毒性の調査は困難であり、AgNPの位置を特定し、ナノコアをAgと区別することが困難になります。私たちのグループは最近、ナノ粒子によって誘発される毒性を研究するための金ナノロッドコア/銀シェル(Au @ Ag NR)ナノ構造を開発しました[17]。 Au @ Ag NRのゴールドコアは組織に生理学的に固有であり、Ag + の放出を監視するための内部標準として使用できます。 誘導結合プラズマ質量分析(ICP-MS)を使用して測定されたAg / Au比の変化を監視することによるロッドからのイオン[18]。この方法により、毒性のさまざまな原因を特定することができます。以前の研究では、放出されたAg + Au @ Ag NRの殻からのイオンは、腎臓の酸化的損傷を引き起こし、最終的には形態学的変化と糸球体の濾過機能の障害を引き起こしました[19]。江ら。 [20]は、Au @ AgNRによる粒子特異的活性と細胞内銀イオン放出の両方が顆粒膜細胞の毒性反応に寄与することを示唆しました。また、AgNPのin vivo遺伝子毒性の可能性を研究するためのモデルとして、Au @ Ag NRを採用し、変異原性ではなく染色体異常誘発性が、Agシェルと放出されたAg + イオン、毒性パターンに違いはありませんでした[21]。
肝臓は、AgNPが蓄積しやすい主要な臓器のひとつであり、AgNPによって誘発される遺伝子毒性の標的臓器/組織として認識されています。私たちの以前の研究では、Au @ Ag NRを1回静脈内投与してから8週間後に、SDラットの肝臓にある程度の銀(8.26±3.90μg/ g)と金(80.07±64.72μg/ g)が残っていることが示されました[21 ]。この研究では、ヒト肝細胞腫由来のHepaRG細胞でAu @ Ag NRを使用して、AgNPによって誘発される染色体およびDNA損傷に対する細胞周期停止および反応性酸化ストレスの役割を特定しようとしました。コメットアッセイ、γ-H2AXアッセイ、微小核試験などの遺伝子毒性アッセイを酸化ラジカルスカベンジャーと並行して実施し、DNA /染色体損傷における反応性酸素種(ROS)の寄与を調べ、細胞アポトーシス、細胞周期、および関連タンパク質を調べました。 AgNPがDNAの合成と複製を妨害するメカニズムを探求することを決意しました。さらに、誘導結合プラズマ質量分析(ICP-MS)と透過型電子顕微鏡(TEM)を組み合わせて、ナノ粒子と放出されたAgイオンの役割を区別することにより、Au @ AgNRの細胞内蓄積と分布を調査しました。
材料と方法
細胞培養と処理
この研究では、ヒト肝癌細胞株HepaRG(Thermo Fisher Scientific)を使用しました。細胞は、5%CO 2 の加湿雰囲気下で、10%ウシ胎児血清(FBS、Australia Origin、Gibco)および1%ペニシリン-ストレプトマイシン-グルタミン溶液(Gibco)を含むRPMI1640で培養しました。 37°Cで。細胞をAu @ Ag NRの濃度を上げながらそれぞれ24時間または72時間処理し、IC 50 に従って濃度を決定しました。 細胞生存率アッセイにより推定。遺伝子毒性におけるROSの役割を調査するには、1 mM N -アセチル-1-システイン(NAC、Sigma-Aldrich)を、Au @ AgNRで処理する前に1時間適用しました。
ATP細胞増殖/生存率アッセイ
細胞を96ウェルプレートに5×10 3 の密度で播種しました。 /良い。 24時間のインキュベーション後、培地を吸引し、細胞をさまざまな濃度のAu @ AgNRにそれぞれ24時間または48時間曝露しました。広範囲の濃度が準備され、1回の処理期間で1回の処理につき4つのウェルが実行されました。 Au @ Ag NRの細胞毒性は、アデノシン三リン酸(ATP)アッセイ(CellTiter-Glo®2.0アッセイ、Promega)によって調べられました。このアッセイは、生細胞の重要な代謝パラメーターであるATPの量を定量することによって細胞の代謝活性を測定します。生細胞の量を反映する発光シグナルは、VICTORマルチラベルプレートリーダー(2030-0050、PerkinElmer)、およびIC 50 を使用して検出されました。 値は、Prism 7(GraphPad Prism 7、CA、USA)によって最大生存率の半分のAu @ AgNRの濃度として推定されました。生存率は、次の式を使用して計算されます。
$$ {\ text {Viability}} \、{\ text {Ratio}} \、\ left(\%\ right)={\ text {RLU}} _ {{{\ text {sample}}}} / { \ text {RLU}} _ {{{\ text {vehicle}}}} \ times {1} 00 \%$$ここで、RLUは、4つのウェルの平均値として表される相対的な光の単位です。RLU Vehicle ナノロッドで処理されていない細胞、およびRLU サンプル さまざまな濃度のAu @ AgNRで処理された細胞を表しています。
セル内の銀と金の濃度測定
細胞サンプルは、マイクロ波分解システムを使用して硝酸で分解されました。分解後、1%硝酸と塩酸を含む混合物でサンプルを調製しました。溶液中のAgとAuの量は、ICP-MS(NexION300X、PerkinElmer)によって決定されました。 TEM分析を使用して、セル内のAuNRおよびAu @ AgNRの存在を確認しました。細胞サンプルを2.5%グルタルアルデヒドと2%パラホルムアルデヒドの混合物で、4°Cで2時間固定しました。細胞ペレットを固定し、リン酸緩衝液(pH 7.4)で3回リンスし、1%四酸化オスミウムで4°Cで2時間後固定しました。続いて、サンプルを蒸留水で3回リンスし、さまざまな濃度のエタノール(それぞれ、50%、70%、90%、100%エタノール)で15分間脱水しました。続いて、1:1および1:3希釈のプロピレンオキシドを、20〜26°Cで2時間樹脂に塗布しました。重合は、35°Cで16時間、45°Cで8時間、55°Cで14時間、65°Cで48時間の段階的加熱によって行いました。超薄切片を酢酸ウラニルとクエン酸鉛で25分間染色し、透過型電子顕微鏡(H-7650、日立、日本)で分析しました。
従来型および修正コメットアッセイ
細胞を2×10 5 の密度で12ウェルプレートに播種しました / wellまたは3×10 5 /ウェルはそれぞれ24時間または72時間の治療に使用します。過酸化水素(H 2 O 2 )200μmolの濃度で、ポジティブコントロールとして1時間細胞に曝露しました。各サンプルについて、従来の処理とホルムアミドピリミジングリコシラーゼ(Fpg)処理の両方のために2つのウェルを準備しました。従来のコメットアッセイは、以前に説明されているようにアルカリ性条件(pH> 13)で実行されました[21]。 Fpg処理ウェルの場合、DNA巻き戻し手順の前に追加のFpg処理を適用し、スライドを酵素バッファー(0.1 M KCl、0.5 mM EDTA、40 mM HEPES、0.2 mg.mL -1)に浸しました。 BSA)各5分間に3回。 Fpg(New England Biolabs、Inc.、UK)を酵素バッファーで1:50,000に希釈しました。希釈した酵素の100ミリリットルのアリコートを顕微鏡のスライド上の各ゲルに加え、37°Cの湿度チャンバーで30分間インキュベートしました。残りの手順は、従来の処理と同じでした。コメットアッセイは3回行った。 Nikon Eclipse 80i蛍光顕微鏡(Nikon、東京、日本)を使用して、サンプルあたり少なくとも50個の細胞を個別にスコアリングし、Komet 6.0(Andor Technology、ベルファスト、英国)を使用して、尾部およびオリーブ尾部のDNAパーセンテージの中間値を分析しました。各サンプルのモーメント(OTM)。
フローサイトメトリーとハイコンテントスクリーニングによるγ-H2AX病巣の認定
フローサイトメトリーを使用した定量化のために、細胞を2×10 5 の密度で12ウェルプレートに播種しました。 / wellまたは3×10 5 /ウェルでそれぞれ24時間または72時間処理し、ハイコンテントスクリーニングアッセイでは、細胞を96ウェルプレートに6×10 3 の密度で播種しました。 / wellまたは1×10 4 /ウェルはそれぞれ24時間または72時間の治療に使用します。ポジティブコントロールとして、2μMのメタンスルホン酸メチル(MMS、Sigma-Aldrich)を細胞と並行して1時間適用しました。細胞をトリス緩衝生理食塩水(TBS)でリンスし、4%パラホルムアルデヒドで室温で15分間固定しました。 TBSで洗浄した後、細胞を50μLの氷冷メタノールとともに-20°Cで30分間インキュベートしました。細胞をさらにTBSで3回リンスし、ブロッキング試薬(0.3%Triton X-100と10%ヤギ血清を含むTBS)を1時間適用しました。一次抗体(マウス抗リン酸化H2AX Ser139、ミリポア)をブロッキング試薬で1:200に希釈し、細胞とともに4℃で一晩インキュベートしました。次に、プレートを再度TBSで3回リンスし、続いてブロッキング試薬で1:20の比率で希釈した二次抗体(Alexa Fluor 488ヤギ抗マウス、Life Technologies)を添加しました。サンプルを室温で1時間暗所に置き、2μgmL -1 (20μL/ウェル)DAPI(Invitrogen)を各ウェルに追加しました。フローサイトメトリー(FACSCalibur、BD Bioscience、ニュージャージー州、米国)またはハイコンテント分析システム(Operetta CLS、PerkinElmer)を使用して蛍光を測定しました。フローサイトメトリーアッセイでは、グループあたり少なくとも10,000個の細胞からのデータを分析し、実験を3回実施しました。ハイコンテント分析では、各ウェルの20の視野と、各グループの少なくとも5つのウェルが分析されました。
細胞質分裂-小核サイトーム(CBMN-cyt)アッセイをブロックする
CBMN-cytは、Fenech etal。によって記述された手順に従って実行されました。 [22]。細胞を2×10 5 の密度で12ウェルプレートに播種しました / wellまたは3×10 5 /ウェルはそれぞれ24時間または72時間の治療に使用します。 0.2μgmL -1 マイトマイシンC(MMC、東京化成工業株式会社、日本)をポジティブコントロールとして24時間細胞に曝露しました。 3μgmL -1 サイトカラシンBは、細胞質分裂プロセスをブロックするために24時間または72時間の処理後に適用され、細胞は40時間後に回収されました。予熱した0.075mol L -1 で低張した後、サンプルを5%ギムザで染色しました。 KClおよびメタノールと酢酸の3:1混合液による固定。グループごとに3つのウェルを準備し、ウェルごとに少なくとも1000個の二核細胞を調べました。
MDA、GSHおよびSODの合計含有量の測定
細胞を12ウェルプレートで5×10 5 の密度で培養しました。 / wellまたは3×10 5 /ウェルはそれぞれ24時間または72時間の治療に使用します。続いて、細胞を回収し、リン酸緩衝生理食塩水(PBS)で3回すすいだ。細胞ホモジネート中のマロンジアルデヒド(MDA)の量は、チオバルビツール酸ベースの方法(Nanjing Jiancheng Bio-engineering Institute、Nanjing、China)を使用して決定されました。総グルタチオン(GSH)とスーパーオキシドジスムターゼ(SOD)の量は、それぞれ総グルタチオン定量化キットとSODアッセイキット(Dojindo Molecular Technologies、Inc。熊本、日本)を使用して決定されました。各ウェルの光学密度(O.D)は、VICTORマルチラベルプレートリーダー(2030-0050、PerkinElmer)を使用して測定しました。
細胞周期のフローサイトメトリー分析
細胞を6ウェルプレートで1×10 6 の密度で培養しました。 / wellまたは5×10 5 /ウェルでそれぞれ24時間または72時間処理し、その後70%エタノールで4°Cで一晩固定しました。サンプルをPBSで3回リンスし、PI / Rnase染色バッファー(BD Biosciences)で室温で15分間染色しました。 20,000個の細胞のうちG0 / G1、S、およびG2 / M期の細胞集団は、FL2面積とFL2幅の領域を使用して決定されました。分析はフローサイトメトリー(FACSCalibur、BD Bioscience、ニュージャージー州、米国)およびFlowJo(BD Bioscience)によって行われ、実験は3回行われました。
細胞アポトーシスのフローサイトメトリー分析
細胞を6ウェルプレートで1×10 6 の密度で培養しました。 / wellまたは5×10 5 /ウェルはそれぞれ24時間または72時間の治療に使用します。続いて、PBSで2回リンスし、500μLの1×結合バッファー(FITCアネキシンVアポトーシス検出キットI、BD Bioscience)で希釈して、懸濁液を約1×10 6 に調整しました。 細胞/ mL、続いて100 µLの希釈液を5 µLのFITCアネキシンVおよび5 µLのPIと混合しました。サンプルを室温で15分間染色し、フローサイトメトリー(FACSCalibur、BD Bioscience、ニュージャージー州、米国)およびFlowJoを使用して、FL1HとFL2Hの領域を使用することにより、アポトーシスの初期および後期の細胞集団を決定するために、少なくとも10,000個の細胞を分析しました。 (BDバイオサイエンス)。実験は3回行った。
ウエスタンブロット分析
細胞は75cm 2 で培養されました 1×10 7 の密度のフラスコ / wellおよび6×10 6 /ウェルはそれぞれ24時間と72時間の治療に使用します。プロテアーゼ阻害剤(PMSF)を含むRIPA溶解バッファーで細胞を溶解し、BCAタンパク質定量キット(Beyotime Biotechnology、中国)を使用してタンパク質の濃度を測定しました。サンプルの濃度は、95°Cで3分間加熱して変性させる前に、RIPA溶解バッファーを使用して調整しました。タンパク質サンプルを12%SDSポリアクリルアミドゲルでの電気泳動により分離し、ニトロセルロースメンブレン(Millipore)に転写しました。メンブレンを5%スキムミルクで30分間ブロックし、一次p53(SC-137174、サンタクルス)、p21(SC-6246、サンタクルス)、β-アクチン(sc-47778、サンタクルス)および二次抗体とインキュベートしました。それぞれヤギ抗マウスIgG(H + L)-HRP(SE131、solabio)。サンプル中の標的タンパク質の発現レベルは、強化化学発光(ECL)法を使用して視覚化され、ImageJシステム(国立衛生研究所)によって分析されました。
統計分析
データは平均±SEMとして表されました。一元配置分散分析(ANOVA)を使用して、陰性対照群と治療群の間の差異の統計的有意性をテストし、続いてSPSS(バージョン22、IBM、アーモンク、ニューヨーク、米国)を使用したダネット多重比較検定を行い、データを検討しました。 P で統計的に有意 <0.05。図は、Windows用のGraphPad Prism 7(GraphPad Software、米国カリフォルニア州ラホーヤ)を使用して作成されました。
結果
AuNRおよびAu @ AgNRの特性評価
金ナノロッド(Au NR)、金ナノロッドコア、および銀シェルナノ構造(Au @ Ag NR)は、前述のように設計、準備、および特性評価されました[21]。簡単に説明すると、平均直径と長さは、AuNRの場合は15.0±2.5nm、66.7±2.5 nm、Au @ AgNRの場合は26.2±3.0nm、72.7±8.9nmです。 Agシェルの厚さは約5nmです。水に分散したPDDACコーティングされたAuNRとAu @ AgNRのゼータ電位はそれぞれ37.7±1.6mVと52.5±1.4mVでした。調製したAu @ AgNRのAg / Au重量比は2.3と推定されました。特性評価の結果を図1に示します。
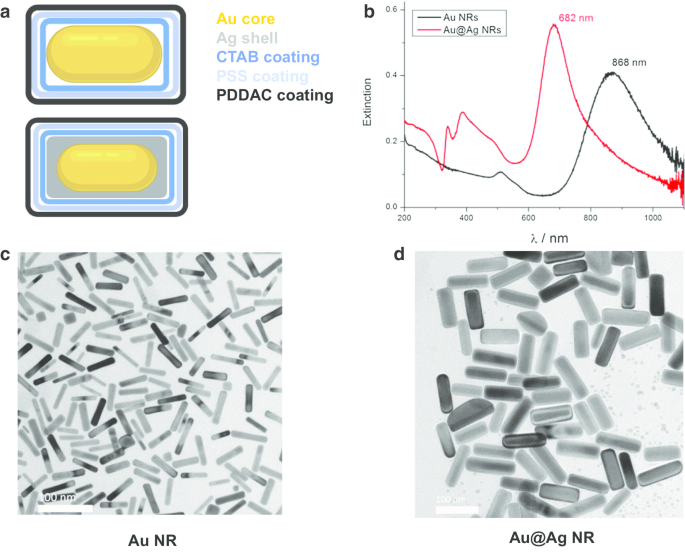
AuNRおよびAu @ AgNRの特性評価。 a AuNRとAu @ AgNRの構造図。 b 水に分散したAuNRおよびAu @ AgNRのUV-Vis-NIR吸光スペクトル。 c AuNRの代表的なTEM画像。 d Au @AgNRの代表的なTEM画像
細胞の生存率
HepaRG細胞に対するAu @ Ag NRの細胞毒性をATP生存率アッセイで調べ(表1)、細胞を0.125〜160μgmL -1の濃度で24時間または48時間Au @ AgNRに曝露しました。 。 Au @ Ag NRは、24時間および48時間の曝露後、時間および用量依存的に有意な細胞毒性効果を誘発し、生存率IC 50 20 µg mL -1 で および6µg mL -1 、ソフトウェアGraphPad Prism7.0によってそれぞれ適合されます。全体的な細胞毒性を考慮して、処理期間を24時間と72時間に調整し、適用した濃度を0.8 µg mL -1 と決定しました。 、4 µg mL -1 および20µg mL -1 。さらに、Au NRは不活性コントロールとして含まれており、AuNRグループのAu含有量は20 µg mL -1 と同じでした。 Au @ Ag NR、16 µg mL -1 。対照的に、Au @ Ag NR + NACグループでは酸化ストレス応答のコントロールとして1mM NAC前処理が採用されました(Au @ AgNRの濃度は20µg mL -1 。
<図>AuNRおよびAu @ AgNRのセル分布
HepaRGセル内のAuおよびAg含有量の分布をICP-MSで分析しました。表2および3に示すように、Ag含有量は用量依存的に増加しました。ただし、抗酸化剤 N -フリーラジカルスカベンジャーとしてのアセチル-l-システイン(NAC)は、同じ濃度のAu @ Ag NR(20 µg mL -1 )でもAg含有量が少ないことが観察されたため、ナノ粒子の細胞取り込みを制限する可能性があります。 )はこのグループに適用されました。 Ag / Au比が24時間から72時間に低下したことは、Ag + が継続的に放出されたことを示しています。 Au @ AgNRのシェルから。また、Agの細胞への取り込みはAuよりはるかに多い(表4)。さらに、TEMデータは、AuNRとAu @ AgNRのほとんどが凝集体としてセル内に保持されていることを示しました。ナノロッドの構造は、核に入らずにAuNRまたはAu @ Ag NRにさらされた細胞の内部にはっきりと見られました(図2)。
<図> <図> <図>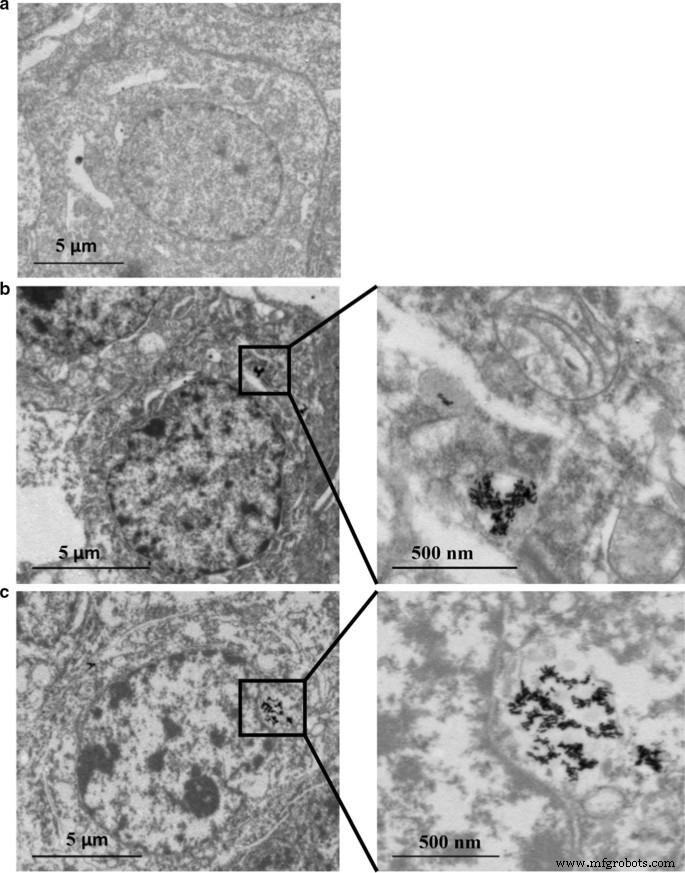
AuNRおよびAu @ Ag NRの内部移行:16μgmL -1 に24時間曝露した後、80kVでTEMによるHepaRG AuNRおよび20μgmL -1 Au @ AgNR。 a 車両制御; b Au NR; c Au @ Ag NR
DNA損傷
Au @ Ag NRによって引き起こされたDNA損傷は、コメットアッセイとγH2AXアッセイの両方で評価されました(図3)。コメットアッセイから、0.8〜20 µg mL -1 Au @ Ag NRは、重大なDNA損傷を引き起こす可能性があります。 Au @ Ag NRへの24時間または72時間の曝露後、細胞の%テールDNAとOTMの両方が、時間依存的および濃度依存的に増加しました。さらに、酸化ストレスの誘導に関連するDNA損傷が、20 µg mL -1 で処理された細胞で観察されました。 Fgp酵素修飾コメットアッセイによるAu @ Ag NR(図3a、b)。癌の発生とのより高い相関を表す二本鎖切断の程度を評価するために、γ-H2AX陽性細胞およびγ-H2AX陽性細胞の平均蛍光強度の両方を分析した。 Au @ Ag NRへの24時間の曝露後、γ-H2AX陽性細胞のグループ間で差は見られませんでした。ただし、4μgmL -1 Au @ Ag NRグループは、72時間の治療後に有意な増加を引き起こしました。ビヒクルコントロールと比較して、72時間後にすべてのAu @ Ag NRグループで蛍光強度の有意な増加が観察されました(図3c–e、 P <0.05)。
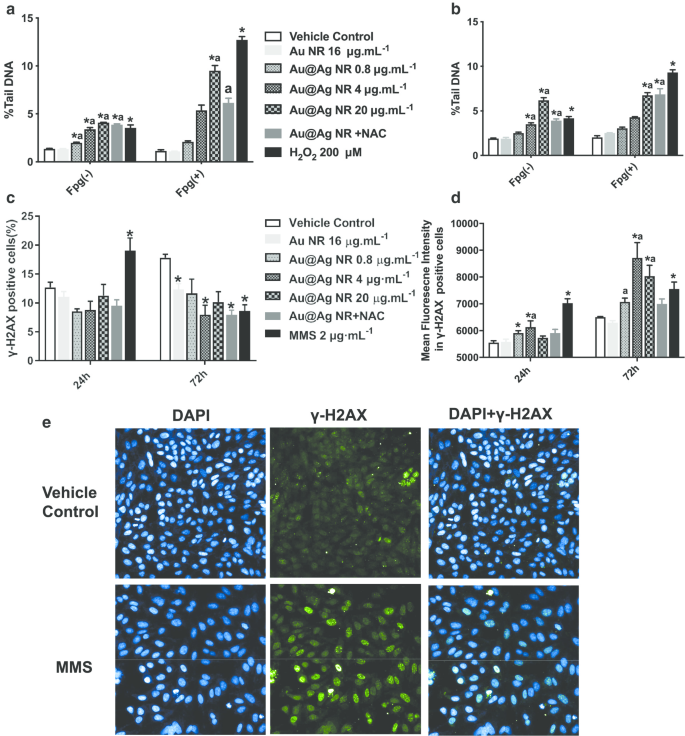
Au @ AgNRによって誘発されるDNA損傷。 HepaGR細胞は、さまざまな濃度(0.8〜20μgmL -1 )のAu @ AgNRに曝露されました。 )それぞれ24時間と72時間。 a Au @ Ag NRに24時間曝露した後の平均%テールDNA。 b Au @ AgNRに72時間曝露した後の平均%テールDNA。 c フローサイトメトリーを使用して推定されたγ-H2AX病巣を有する陽性細胞のパーセンテージ。 d 免疫蛍光染色を使用して推定されたγ-H2AX病巣を有する細胞の平均蛍光強度。 * P <0.05対車両制御; a P <0.05対AuNR。 2μMmL -1 MMSをポジティブコントロールとして採用しました
染色体損傷
小核の形成は染色体損傷を特定するための重要なバイオマーカーであり、これはDNA破壊よりも遺伝物質へのより重大な損傷です。小核を含む二核細胞の比率は、図4cに示すようにスコアリングされました。 Au @ Ag NRは、濃度依存パターンで小核形成を増加させました。 24時間の曝露後、4μgmL -1 で処理された細胞で観察された小核の比率 Au @ AgNRおよび20μgmL -1 Au @ Ag NRはそれぞれ1.133±0.145%と1.567±0.318%であり、どちらもビヒクル対照群よりも有意に高かった。 72時間の曝露後、4μgmL -1 で処理された細胞の小核の比率 Au @ Ag NRは1.767±0.233%であり、これはビヒクル対照群よりも有意に高かった。 20μgmL -1 で処理された細胞の小核の比率 Au @ Ag NRは2.167±0.252%であり、ビヒクル対照群と16μgmL -1 の両方で観察されたものよりも有意に高かった。 Au NRグループ(0.700±0.153%)。対照的に、20μgmL -1 で処理した細胞間には違いは見られませんでした。 Au @ Ag NR + NACおよびビヒクル制御。これは、Au @ AgNRによって誘発される染色体破壊へのROSの関与を示唆しています。
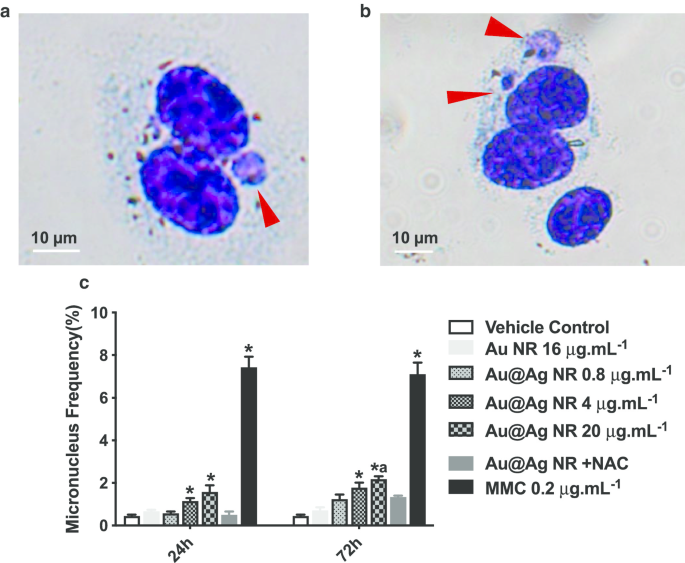
Au @ AgNRによって誘発される染色体損傷。 HepaGR細胞は、0.8μgmL -1 とは異なる濃度のAu @ AgNRに曝露されました。 〜20μgmL -1 24時間と72時間。 a 、 b 小核の代表的な画像(赤い矢印); c 小核頻度(%)。 * P <0.05対車両制御; a P <0.05対AuNR。 0.2μgmL -1 マイトマイシンCを陽性対照として使用しました
ROS形成に対するAu @ AgNRの影響
Au @ Ag NRによって誘発されるDNAおよび染色体の損傷におけるROS形成の役割をさらに調査するために、MDA、GSH、およびSODレベルを推定しました。 MDA形成の大幅な増加( P <0.05)は20μgmL -1 への曝露後に観察されました 24時間と72時間の両方のAu @ Ag NR(図5a)。さらに、Au @ Ag NRに曝露された細胞のGSHおよびSODレベルは、有意な減少を示しました( P <0.05)時間および濃度に依存する方法で。これらの結果は、Au @ Ag NRの曝露によって生成された、酸化と抗酸化の不均衡を示唆しています(図5b、c)。
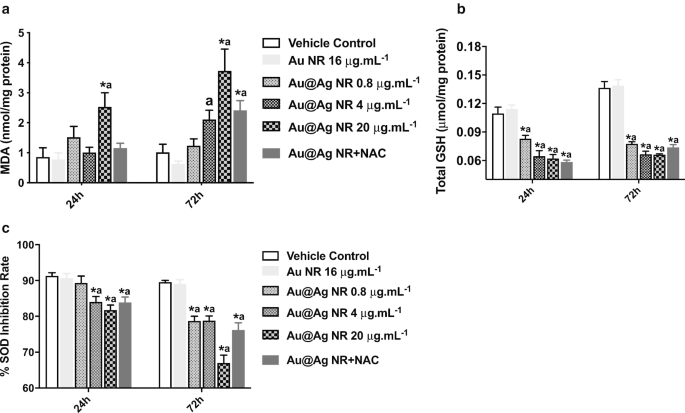
ROS形成に対するAu @ AgNRの影響。 HepaGR細胞は、0.8μgmL -1 とは異なる濃度のAu @ AgNRに曝露されました。 〜20μgmL -1 24時間と72時間。 a MDAレベル; b GSHレベル; c SODレベル。 * P <0.05対車両制御; a P <0.05対AuNR
細胞周期とアポトーシスに対するAu @ AgNRの影響
Au @ Ag NRに72時間曝露した後、G2 / M期の細胞数の増加が4μgmL -1 で観察されました。 Au @ Ag NR、20μgmL -1 Au @ AgNRおよびAu @ Ag NR + NACグループは、それぞれ32.63%±1.77%、32.267%±2.17%および32.967%±4.25%の割合であり(図6a、b)、これらは有意に大きかった。車両対照群(22.37%±0.92%)。一方、Au @ Ag NRによって誘導される細胞アポトーシスは、72時間の曝露後に観察でき、20μgmL -1 で処理された細胞のアポトーシス後期率が観察されました。 Au @ AgNRおよび20μgmL -1 Au @ Ag NR + NACはそれぞれ78.90±1.19%と70.20±4.50%でした(図6c、d)。 Au @ Ag NRは初期アポトーシスよりも後期アポトーシスを誘発し、NACの治療は、Au @ AgNRによって引き起こされる後期アポトーシスの細胞率を軽減する可能性があります。
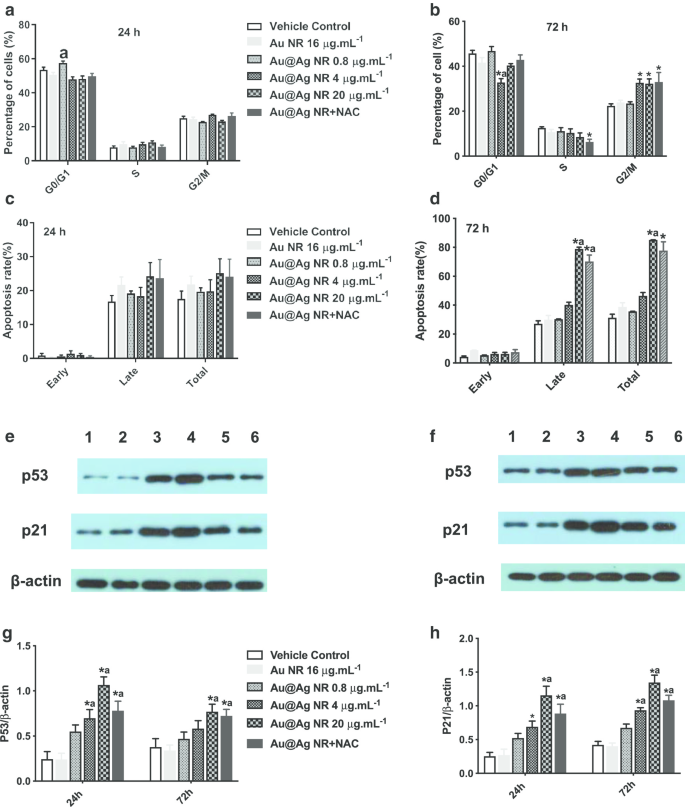
細胞周期とアポトーシスに対するAu @ AgNRの影響。細胞周期に対するAu @ Ag NRの影響( a 、 b )およびアポトーシス( c 、 d )それぞれ24時間と72時間曝露した後。異なるグループのHepaRG細胞におけるp53およびp21の発現レベルの代表的なデータ( e 、 f レーン1:車両制御;レーン2:Au NR;レーン3:Au @ Ag NR + NAC;レーン4:Au @AgNR20μgmL -1 ;レーン5:Au @AgNR4μgmL -1 ;レーン6:Au @AgNR0.8μgmL -1 );異なるグループにおけるβ-アクチンに対するp53およびp21の平均相対発現レベルを( g 、 f )。 * P <0.05対車両制御; a P <0.05対AuNR
p21とp53の発現レベルはウエスタンブロットで検出され、同様のパターンが観察されました。 4μgmL -1 で処理した細胞におけるp53とp21の発現レベル および20μgmL -1 Au @ Ag NRは著しく増加しました( P <0.05)、20μgmL -1 の両方で処理した細胞で有意に減少しました Au @ Ag NRおよびNAC( P <0.05、20μgmL -1 と比較 Au @ Ag NRグループ、図6e–h)。 p53タンパク質はDNA損傷に応答してG2 / Mチェックポイントの活性化を媒介するコア分子であることが知られており、p21はp53依存性の細胞周期阻害剤として認識されています。したがって、Au @ Ag NRはDNA複製を妨害し、細胞周期停止によるDNA修復を妨げる可能性があります。
ディスカッション
現在、リリースされたAg + の役割 遺伝子毒性の発生におけるAgNPは明らかではありません。私たちのグループ[21]および他のグループ[13]による以前の研究では、Ag + 毒性を導入する主な原因であるナノ粒子は、非常に毒性が高い可能性もあります。たとえば、AgNPは、ヒドロキシルラジカルの形成を誘発することによって遺伝子毒性に寄与する可能性があります[13]。さらに、Ag + と比較して、AgNPによって、より深刻な染色体損傷、酸化ストレス、およびアポトーシスが導入されました。 単独で[23]、異なる経路が関与している可能性があることを示唆している。細胞内のAgNPの形態と分布を理解するためのモデル材料としてAu @ Ag NRを採用し、細胞内のAgとAuの量をICP-MSで測定しました。調製したAu @ AgNRのAg / Au重量比は2.3と推定されました。 However, after a 24-h exposure, it sharply increased to 16.5 in the cells treated with Au@Ag NR, suggesting that large amount of Ag was released from the shell of Au@Ag NR within that period. When the exposure period of Au@Ag NR was extended to 72 h, the Au/Ag weight ratio was decreased to 1.7, indicating that the Ag + was released from the cell and the nanorod was the major form of Au@Ag NR in the cell at that stage. Therefore, it could be deduced that once the Au@Ag NR entered the cell, Ag + rapidly dissolved from its shell within 24 h and gradually released to the extracellular environment, while the Au@Ag NR itself retained in the cell for a longer period.
Oxidative stress is deemed as one of the most important toxicological mechanisms of nanoparticles [24]. N-acetylcysteine (NAC) is a thiol, a mucolytic agent and precursor of l-cysteine which reduced glutathione. NAC is also a source of sulfhydryl groups in cells and exerts the ROS scavenger activity by interacting with OH · およびH 2 O 2 [25]。 In this study, the GSH and SOD levels were significantly decreased after exposure to Au@Ag NR, while the MDA level increased in a concentration- and time-dependent manner, indicating that the Au@Ag NR introduced the oxidative stress in the cells.
The potentials of Ag and Au@Ag NR in interfering with the genetic materials were further investigated by a series of genotoxicity assays. It is noteworthy that co-culturing the NAC with Au@Ag NR could ameliorate the ROS formation, which in turn supports the participation of oxidative stress in the genotoxicity triggered by Au@Ag NR. In this study, comet and γ-H2AX assays were performed to confirm that Au@Ag NR could interact with DNA and induce certain DNA damage, and the repair endonuclease Fpg was included in the comet assay to identify the oxidative DNA damage [26]. The Fgp could recognize oxidized pyrimidines and remove oxidized purines, e.g., 8-hydroguanine, so as to create apurinic or apyrimidinic sites that could introduce gaps in the DNA strands. The oxidative stress-induced DNA breakage could be determined subsequently by another comet assay [27]. The further DNA breakage detected by the additional Fgp in the comet assay suggested that the Au@Ag NR could cause DNA damage. Mei et al. [28] observed that 5-nm-sized AgNPs induced oxidative lesion-specific DNA damage by employing the hOGG1, EndoIII and Fpg endonucleases in the comet assay. Li etal。 [29] also suggested that both PVP- and silica-coated AgNPs (15–100 nm and 10–80 nm, respectively) could lead to a significant increase in DNA breakage in mice hepatocytes in the presence of hOGG1and EndoIII. The formation of γ-H2AX foci, which represents an early cellular response to genotoxic stress, is the most sensitive and specific biomarker for detecting DSBs [30]. As demonstrated in this study, γ-H2AX foci in cells exposed to Au@Ag NR were markedly increased after 24 h, and a further increase could be observed after 72 h. The reduction in the 20 µg mL −1 group might be due to the cytotoxicity to the HepaRG cells at higher concentration. Similar results were observed for AgNPs with different coatings [31, 32]. Further, our results suggest that Au@Ag NR could induce chromosome damage in HepaRG cells, as the micronucleus rates were significantly increased. This is consistent with previous studies, where AgNPs-induced increased micronucleus rate was reported in HaCaT and TK6 cells [33]. In contrast, the addition of oxidative radical scavenger NAC could inhibit the formation of micronucleus induced by Au@Ag NR. Taken together, these data suggest the participation of oxidative stress in AgNP-introduced clastogenicity risk in vitro.
Previous studies have investigated the cell cycle arrest and cytotoxicity induced by AgNPs [33,34,35]. With prolonging the exposure time, the impact of AgNPs on cell cycle and apoptosis might be enhanced and in turn aggravate the cytotoxicity and genotoxicity. Usually, the cell cycle checkpoints (e.g., G2/M) were initiated by cells when experiencing DNA damage, and this mechanism serves to prevent the cell from entering mitosis (M phase). The G2/M cell cycle arrest indicates that an increasing percentage of cells is hindered in G2 phase for DNA repairing. Cells experiencing successful DNA repairing would further proceed to mitosis; however, for those with fatal damages, irreversible G2/M cell cycle arrest and cells apoptosis would take place [36]. We observed that Au@Ag NR could arrest the majority of HepaRG cells in G2/M phase, induce late cell apoptosis and increase the expression levels of p53 and p21, which are important proteins associated with the regulation of cell cycles [37]. As p53 could also induce apoptosis, when the DNA cannot be repaired properly [38], the p21 might indirectly participate in cell apoptosis by cell cycle arrest in a p53-dependent pathway via down-regulating the nuclear protein ICBP90 for DNA replication and cell cycle regulation [39]. Furthermore, apoptosis and a G2/M arrest induced by activation of the p53/p21 system have been reported in HepG2 cells following the administration of garlic extracts [40]. Thus, it could be inferred that the oxidative stress-triggered DNA/chromosome damages might facilitate the expression of p53 and p21, which subsequently induces cell cycle arrest. Extending the exposure period of Au@Ag NRs to the DNA/chromosome during replication may further aggravate the genotoxicity or apoptosis.
結論
Genotoxicity induced by AgNPs may be attributed to the oxidative stress induced by the nanoparticles as well as the released ions [41]. This study employed Au@Ag NR as a model to determine the distribution and release behavior of Ag after the nanoparticles enter into the cells. Considering the disparate forms of Au@Ag NR in the cell, after its exposure the Ag + was rapidly dissolved from the silver shell. Ag + and Au@Ag NR could introduce cytotoxicity and genotoxicity (clastogenicity) in the cells, and the Au@Ag NR retained in the nucleus may further release Ag + to aggravate the damage, which are mainly caused by cell cycle arrest and ROS formation (summarized in Fig. 7). Collectively, these data reveal the correlation between the intracellular accumulation, Ag + release as well as the potential genotoxicity of AgNPs.

Schematic diagram of the possible mechanism of genotoxicity introduced by AgNP in vitro
データと資料の可用性
All data and materials are available without restriction.
ナノマテリアル
- タングステン銀合金とタングステン銅合金の違い
- ICAをロードしたmPEG-ICAナノ粒子の調製とLPS誘発性H9c2細胞損傷の治療におけるそれらの応用
- ミセルナノ結晶の界面不安定性に基づく製造プロセスにおけるエマルジョン液滴サイズと界面活性剤の役割の検討
- 色素増感太陽電池の光学的および電気的特性に及ぼすTiO2中の金ナノ粒子分布の影響
- Au @ TiO2卵黄シェルナノ構造の調製とメチレンブルーの分解および検出へのその応用
- 6-メルカプトプリンとニューロン透過性ペプチドで修飾された金ナノ粒子によるSH-SY5Y細胞増殖の促進
- CoFe2O4 / Fe3O4およびFe3O4 / CoFe2O4コア/シェルナノ粒子における深い界面効果
- 金および銀ナノ粒子のグリーン合成のためのPlatycodiRadix(Platycodon grandiflorum)からのPlatycodonサポニン
- Cryptococcusneoformansの診断と光熱治療のための抗体結合シリカ修飾金ナノロッド:invitroでの実験
- 食用菌根菌Tricholomacrassumから合成されたタンパク質でキャップされた金ナノ粒子の抗菌性、アポトーシス性、および癌細胞の遺伝子送達特性の評価
- CdフリーのCuドープZnInS / ZnSコア/シェルナノ結晶:制御された合成と光物理的特性