


アルテスナートのナノ粒子送達は、ミトコンドリアを介した細胞アポトーシスを活性化することにより、抗腫瘍効率を高めます
要約
アルテミシニンとその誘導体は、広範囲の抗癌活性を発揮すると考えられており、腫瘍細胞に有意な抗癌効果を誘発しました。アルテミシニンとその誘導体は迅速に吸収され、広く分布し、腫瘍細胞を選択的に殺しました。低濃度のアルテスナートは主に腫瘍細胞の細胞死を誘発するために腫瘍症に依存していたため、その抗腫瘍効果は望ましくなく、制限されていました。より優れた抗腫瘍効果を得るために、この研究では、新しいナノテクノロジーを利用して、アルテスナートのミトコンドリア蓄積を達成し、ミトコンドリアを介したアポトーシスを誘導する、アルテスナートをロードした新しいウシ血清アルブミンナノ粒子を設計しました。結果は、遊離アルテスナートの膠質浸透圧死への依存と比較した場合、アルテスナートをロードしたウシ血清アルブミンナノ粒子はより高い細胞毒性を示し、ミトコンドリア内のアルテスナートの分布を通じてそれらの有意なアポトーシス効果が誘導されたことを示しました。この発見は、アルテスナートをロードしたウシ血清アルブミンナノ粒子が、アポトーシス関連タンパク質をアップレギュレートし、シトクロムCの急速な放出を促進することにより、ミトコンドリアの完全性を損傷し、ミトコンドリアを介した細胞アポトーシスを活性化したことを示しています。
背景
アルテミシニンとその誘導体は、抗マラリア活性が高く、毒性が低いため、マラリアの治療に広く使用されています。研究者はまた、アルテミシニンとその誘導体が、それらのわずかな毒性副作用と患者によるより大きな耐性のおかげで、有意な抗腫瘍活性を示したことを発見しました[1]。アルテスナート(Ats)は腫瘍細胞の成長を確実に阻害し、腫瘍細胞に有意な抗癌効果をさらに誘発することが報告されました[2、3、4]。一部の実験では、Atsが48時間後に腫瘍細胞にさまざまな程度のアポトーシスと腫瘍症を引き起こし、アポトーシスと腫瘍症の程度はAtsの用量に依存することが示されました。低濃度では、Atsは腫瘍細胞に明らかなアポトーシスを誘発せず、Ats誘発細胞死は腫瘍症様死を伴っていました[5,6,7,8]。より大きな抗腫瘍効果を得るために、より高い投与量のAtsが適用されましたが、これにより、その深刻な毒性と骨髄抑制がさらに確認されました。したがって、抗腫瘍効果を高めるために、Atsの有効量を減らすための効果的な治療法を見つける必要があります[9、10、11]。ミトコンドリアは、Atsのアポトーシスおよび膠質浸透圧効果の調節に重要な役割を果たしていることがわかりました。ミトコンドリアは、多種多様なアポトーシスシグナルの伝達プロセスの調節にも関与していました[12、13、14、15、16、17]。ミトコンドリアが薬物によって攻撃されると、その透過性が高まり、膜電位が低下したため、ミトコンドリア膜の子宮内膜の腫れと、ミトコンドリアから細胞質へのシトクロムCの急速な放出が起こりました[18、19、20]。さらに、カスパーゼファミリーの一部のタンパク質が活性化され、細胞アポトーシスのカスケード反応が誘導されました。
Atsの抗腫瘍効果を高めるために、腫瘍細胞における薬物の分布を増加させたり、細胞小器官への薬物の標的送達を改善して細胞死を誘発したりするために、多くの新しい技術が試みられました[21、22、23]。標的癌治療の重要なツールとしてのナノ粒子(NP)は広く研究されており、有望な可能性を示しています。 NPは粒子サイズが小さく、表面積が大きいため、毛細血管を介して血液循環に入り、内皮細胞のギャップを通過して腫瘍部位に移動し、薬物を標的とした分布を実現し、薬物のバイオアベイラビリティを高めます。 。さらに、NPは、生体材料を長く滑らかなパターンで分解することにより、薬物の放出を制御し、最終的には半減期を延長し、有効血中濃度を改善し、投与頻度を減らすことができます。何よりも、薬物をロードしたNPを細胞内の特定の場所に送達して、治療効果を向上させることができます[24、25、26]。
低濃度でのAtsの抗腫瘍効果を高めるために、新しいAtsをロードしたウシ血清アルブミン(BSA)NPの設計を試みました。腫瘍細胞のpHが低いため、ミトコンドリア外膜または膜間腔に多数の水素プロトンが蓄積します。逆に、ミトコンドリア間膜は、その化学組成とミトコンドリアマトリックスのために負の電荷が豊富です。分泌、それはBSAの有利な送達をすることができる膜電位の外側と負の電気陽性を作ります。次に、ミトコンドリアにおけるAtsの大量の蓄積は、ミトコンドリアを介したアポトーシスを効果的に誘発する可能性があります。結果は、遊離Atsによって誘発される典型的な膠質浸透圧死と比較した場合、AtsはBSA NPを介してミトコンドリアに特異的に移動し、ミトコンドリアを介したアポトーシス関連カスパーゼタンパク質の活性化を促進することを示しました。これは有意な細胞アポトーシスを引き起こし、したがってより高い細胞毒性を強調しました。
メソッド
資料
BSAはSigma-AldrichCo。(セントルイス、ミズーリ州、米国)から購入し、AtsはGuilin Pharmaceutical Corporation(桂林、中華人民共和国)から購入しました。 SMMC-7721細胞およびPlc細胞は、中国科学院(上海、中華人民共和国)の生化学および細胞生物学研究所から購入しました。購入した他のすべての化学物質は分析グレードのものでした。さまざまなベンダーから入手しました。
AtsをロードしたBSANPの準備と特性評価
以前に報告された文献[27]によると、AtsをロードしたBSANPは脱溶媒和法によって調製されました。簡単に説明すると、AtsをロードしたBSA NPは、一定量のAtsを含む1.0mLの無水アルコールを37°Cで乳白色になるまで0.5mLのBSA溶液にすばやく滴下することによって調製しました。回転蒸発によるエタノールの除去に伴い、AtsをロードしたBSA NPを培地からさらに沈殿させ、次に水中の8%グルタルアルデヒド(0.5μL/ mgBSA)を添加して、懸濁液を一定時間攪拌しながら粒子の架橋を誘導しました。 24時間の最後に、NPを収集し、脱イオン水で3回洗浄して、流体力学的直径、多分散性指数(PDI)、ゼータ電位、ブルックヘブンゼータサイザー(ブルックヘブンインスツルメンツコーポレーション、ニューヨーク州ホルツビル、米国)を使用した形態などの物理的特性をさらに分析しました。および透過型電子顕微鏡(JEM-1200EX; JEOL、東京、日本)。 BSA NPにおけるAtsのカプセル化効率の決定は、以前に報告された方法[27]を使用して推定されました。
MTTアッセイ
SMMC-7721細胞とPlc細胞の2種類の腫瘍細胞株を、20%ウシ胎児血清(FBS)と別々にインキュベートしました。細胞増殖密度をl×l0 6 に調整しました 細胞数で細胞/ mL、次に細胞懸濁液をl×l0 5 に希釈しました 細胞/ mL。希釈した懸濁液をさらに別々に96ウェルプレートに加えました(ウェルあたり100μL、約1×10 4 細胞/ウェル)5%CO 2 の条件下で、37°Cで24時間の連続インキュベーション および95%O 2 。培地を、遊離のAtsまたは異なる濃度のAtsを特徴とするAtsをロードしたBSA NPのいずれかの存在下で無血清培地に交換し、その後24時間インキュベートしました。合計50μLの3-(4,5-ジメチルチアゾール-2-イル)-2,5-ジフェニルテトラゾリウムブロミド(MTT)(5 mg / mL)を各ウェルに添加し、培養を終了するために4時間インキュベートしました。テトラゾリウム色素MTTが不溶性のホルマザンに還元されたら、96ウェルプレートを1000 rpmで5分間遠心分離し、各ウェルから上清をデカントした後、150μLのジメチルスルホキシド(DMSO)を添加しました。結晶を溶解しました。溶液の吸光度は、マイクロプレートリーダー(Syneray-2; BioTek Instruments、Inc.、Winooski、VT、USA)を使用して490nmで測定しました。
細胞内のBSANPグループの細胞内分布
対数期のSMMC-7721細胞とPlc細胞を選択し、トリプシン消化で処理しました。細胞濃度をl×l0 6 に調整しました 細胞/ mL。次に、培養細胞を接着のために6ウェル細胞培養プレートに加え、培養培地を除去した後、ローダミンB標識BSANPを加えた。核はヘキスト(青)で37°Cで15分間染色され、ミトコンドリアはMitotracker GreenFMで染色されました。細胞内のBSANPの位置は、共焦点レーザー走査顕微鏡(FluoView FV10i;オリンパスコーポレーション、東京、日本)を使用して細胞内で追跡されました。
ミトコンドリア膜電位の変化
JC-1は、ミトコンドリア膜電位の変化を測定するために使用できます。ミトコンドリア膜電位が高い場合、JC-1は細胞膜を自由に通過し、ミトコンドリア内に凝集体を形成し、赤色蛍光を示しました(励起波長525 nm、発光波長590 nm)。ミトコンドリア膜電位が低下すると、JC-1はミトコンドリアマトリックスから細胞質に移動し、緑色の蛍光モノマー(励起波長、490 nm、発光波長、530 nm)を形成します。 SMMC-7721細胞とPlc細胞をそれぞれ共焦点ディッシュに播種し、密度がl×l0 6 になるようにしました。 12時間の連続インキュベーションの場合はcells / mL。次に、培地を廃棄し、AtsまたはAtsをロードしたBSANPの分散液を含む無血清培地をディッシュに加えました。 9時間後、培地を廃棄し、細胞をPBSで2回洗浄した後、2μmol/ Lの濃度で2mLのJC-1を添加しました。次に、細胞を暗所で37℃で30分間インキュベートしました。レーザー走査型共焦点顕微鏡(FluoView FV10i; Olympus Corporation)を使用して、ミトコンドリア膜の画像変化を観察しました。
小胞体(ER)のROS産生測定と染色
細胞を20%FBSでインキュベートし、細胞増殖密度を1×10 6 に調整しました。 細胞数による細胞/ mL;次に、細胞懸濁液を1×10 5 に希釈しました。 細胞/ mL。希釈した懸濁液をさらに96ウェルプレートに加えました(ウェルあたり100μL、約1×10 4 細胞/ウェル)5%CO 2 の下で37°Cで24時間の連続インキュベーション および95%O 2 。次に、遊離のAtsおよびAtsをロードしたBSA NPを細胞と6、12、および24時間インキュベートした後、10μMの2,7-ジクロロフルオレセインジアセテート(DCFH-DA; Sigma-Aldrich Co.)と連続的にインキュベートしました。約30分。氷冷PBSバッファーを使用して細胞を3回洗浄し、内在化されていないNPを除去しました。 485 nmで励起され、530 nmで放出される細胞内DCF蛍光強度は、酸化ストレスの程度を調査するためにマイクロプレートリーダー(Synergy-2; BioTek Instruments)を使用して検出されました。テストグループをSMMC-7721セルとPlcセルで24時間処理し、ER-Tracker Blue–White DPXプローブ(Molecular Probes、米国オレゴン州ユージーン)をセルに追加して30分間インキュベートしました。ローディング溶液を廃棄し、細胞をPBSで洗浄した後、共焦点レーザー走査顕微鏡でERの形態変化を観察しました。
フローサイトメトリーによる細胞死とアポトーシスの評価
以前の研究のプロトコル[28]によると、アネキシンV-フルオレセインイソチオシアネート(FITC)/ヨウ化プロピジウム(PI)染色アッセイを使用して、遊離のAtsおよびAtsをロードしたBSANPによって誘導される細胞の発癌およびアポトーシスを評価しました。細胞をタイプシンで溶解し、l×l0 6 の濃度で6ウェルプレートに播種しました。 24時間の連続インキュベーションで細胞/ mL。次に、培地を除去し、遊離のAtsおよびAtsをロードしたBSANPを含む無血清培地をウェルに加えた。処理後、細胞を収集し、PIおよびFITC標識アネキシンV(AV-FITC)を含むニコレッティバッファー(北京4Aバイオテック株式会社、北京、中華人民共和国)に懸濁しました。細胞の形態学的変化は、共焦点レーザー走査顕微鏡によって観察された。 AtsをロードしたNPによって誘導される細胞アポトーシスと発癌率を検証するために、初期アポトーシス(Q4)、発癌(Q2)、壊死(Q1)、および生細胞(Q3)のパーセンテージをフローサイトメトリーで定量化しました。
>細胞内のアポトーシス関連タンパク質とシトクロムCのウエスタンブロット分析
ウエスタンブロットアッセイを実施して、遊離のAtsまたはAtsをロードしたNPをSMMC-7721細胞と24時間インキュベートしたときの相対タンパク質のレベルを決定しました。プロテアーゼ阻害剤カクテルとホスファターゼ阻害剤(Roche、Basel、Switzerland)を含む氷冷放射免疫沈降アッセイ(RIPA)バッファーで細胞を溶解しました。タンパク質濃度は、修正BSAアッセイキット(Thermo Fisher Scientific、マサチューセッツ州ウォルサム、米国)を使用して決定し、10%ドデシル硫酸ナトリウム(SDS)-ポリアクリルアミドゲル電気泳動(PAGE)にロードする前に正規化しました。標的タンパク質のレベルを写真に撮り、UVPゲル分析システム(iBox Scientia 600; UVP、LLC。、Upland、CA、USA)を使用して分析しました。
結果
AtsをロードしたBSANPの特性と細胞生存率の研究
図1a、bで、AtsをロードしたBSA NPは球形を示し、0.016の低いPDIで均一に分散していることが観察されました。 AtsをロードしたBSANPの平均粒子サイズは約99.9±2.3nmで、ゼータ電位は負であり、約–25.6±4.3mVと評価されました。放出プロファイルは、図1cに示すように、スムーズで持続的な放出を示しました。インビトロでの培地における遊離Atの急速な放出と比較して、BSA NPのコアに捕捉されたAtは、NPの内部から培地にゆっくりと拡散し、BSAの継続的な分解により、滑らかで持続的な放出パターンを示した。無料のAtの85%以上が、最初の6時間以内に完全に放出されましたが、48時間以内にNPからメディアに放出された薬物の総累積量は78.9%でした。これは、NPが生体材料の分解を介して薬物の放出を長く滑らかなパターンで制御できることを示しています。これにより、排出半減期が延長され、有効血中濃度が改善され、投与頻度が減少します。
<画像>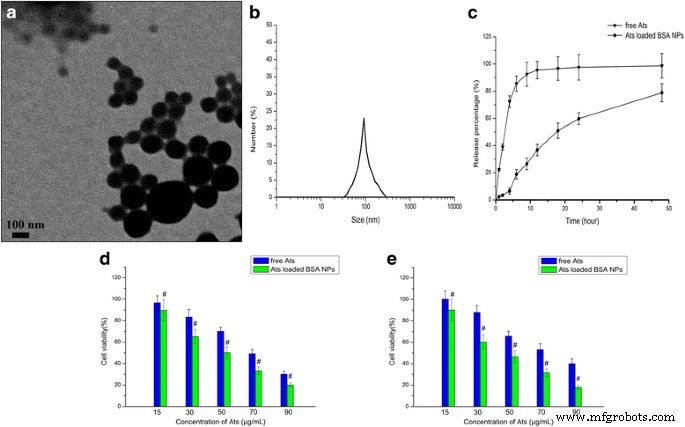
AtsをロードしたBSANPの特性評価。 a AtsをロードしたBSANPのTEM画像。 b 得られたAts負荷BSANPの動的光散乱(DLS)分析。 c AtsをロードしたBSANPのリン酸緩衝生理食塩水(pH 7.4、37°C、48時間)でのinvitro放出プロファイル。 SMMC-7721細胞の生存率( d )およびPlcセル( e )さまざまな量の無料のAtsおよびAtsをロードしたBSANPと24時間インキュベートした後。データは平均±SD( n )として表されます =3)。 # P <0.05対対応する無料Ats
MTTを使用して、SMMC-7721細胞およびPlc細胞における遊離AtsおよびAts負荷BSANPの阻害効果をさまざまな時間間隔で調べました。結果(図1d、e)は、遊離Atsの細胞毒性が薬物濃度の増加とともに増加し、AtsをロードしたBSANPが徐々に細胞毒性を増強することを示しました。これは、AtsおよびAtsをロードしたBSA NPが腫瘍細胞の増殖を阻害し、阻害率がAtsの用量に依存することを証明しました。遊離のAtと比較して、AtsをロードしたBSA NPは、両方の細胞でより高い細胞毒性とより高い感度を示し、より大きな細胞阻害をもたらしました。図1d、eに示すように、AtsをロードしたBSA NPで両方の細胞を処理すると、遊離のAtsと比較した場合、24時間で細胞生存率が大幅に低下しました。 AtsをロードしたBSANPで処理したSMMC-7721細胞とPlc細胞の50%最大阻害濃度(IC50)値は、24時間でそれぞれ50.1と44.9μg/ mLであり、69.2と74.9の値と比較されます。遊離Atsで処理された細胞における24時間でのμg/ mL。これは、AtsがBSA NPにロードされると、NPによって媒介されるように、細胞内の位置を変更し、最終的にはより多くの細胞を殺す可能性があることを示しています。
BSANPのinvitro細胞取り込み
図2に示すように、両方のタイプの腫瘍細胞におけるBSA NPの細胞内分布と位置を、共焦点レーザー走査顕微鏡で観察しました。ローダミンB標識NPを細胞と3時間共培養した後、赤色の蛍光がはっきりと見られました。細胞質内;時間の経過とともに、BSA NPの大部分は細胞内に内在化され、細胞質に拡散し、時間依存性の赤色蛍光が増強されました。黄色の蛍光の出現によって明らかなように、細胞質に位置するBSA NPがミトコンドリアと同じ場所にあることも観察されました。これは、ローダミンBで標識されたNPの固有の赤色蛍光とミトコンドリアインジケーターMitoTracker®緑色FMが統合されました。これは、内在化されたBSA NPがミトコンドリア内に特異的に蓄積される可能性があることを証明し、BSANPの媒介によりAtsがミトコンドリアに送達される可能性を浮き彫りにしました。
<画像>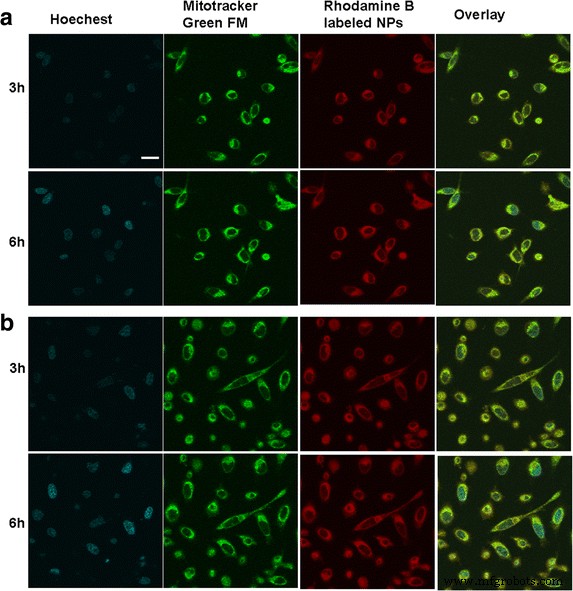
異なる腫瘍細胞とインキュベートした後のBSANPのinvitro細胞分布。 SMMC-7721細胞の蛍光画像( a )およびPlcセル( b )。 スケールバー 、100μm
ミトコンドリア膜電位分析
AtsをロードしたBSANPがミトコンドリアでのAtsの送達に続いてミトコンドリア機能を妨害したかどうかを明らかにするために、ミトコンドリア膜電位の変化を測定しました。図3は、JC-1染色後、遊離Atsで処理された腫瘍細胞のミトコンドリアの大部分が強い赤色蛍光と弱い緑色蛍光強度を示したことを示しています。これは、JC-1の大部分が凝集状態で存在し、ミトコンドリア膜の完全性とより高い可能性を強化していることを示唆しました。逆に、JC-1がAtsをロードしたBSA NPで処理した細胞を染色すると、両方の腫瘍細胞のミトコンドリアがより強い緑色の蛍光を示し、ミトコンドリア膜が深刻に損傷し、その電位が大幅に低下したことを示しています。まとめると、AtsがBSA NPを介してミトコンドリアに正常に送達され、ミトコンドリア膜の脱分極が生じることが証明されました。
<画像>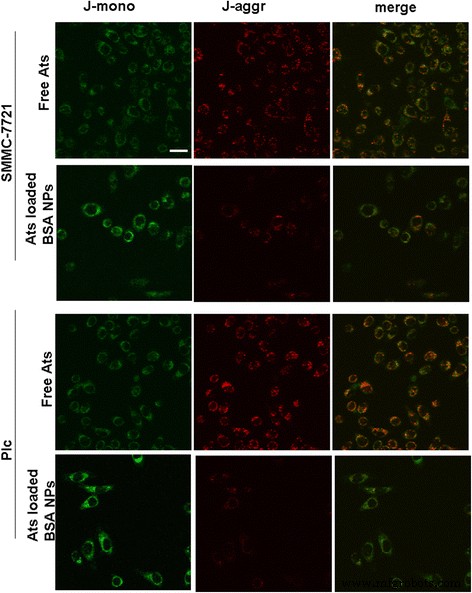
遊離AtsおよびAtsをロードしたBSANPをSMMC-7721細胞およびPlc細胞とインキュベートした後のミトコンドリア膜電位の画像変化。 スケールバー 、100μm
ROS生産測定とERの染色
多数のROSが生成されると、ミトコンドリア内膜でリン脂質の過酸化が起こり、ミトコンドリア膜電位が低下し、シトクロムCが急速に放出されることが広く確認されています。DCFH-を使用しました。 ROSの変化を検出するための蛍光プローブとしてのDA。 DCFH-DAは細胞膜を自由に通過して細胞に入り、エステラーゼ加水分解によってDCFHに変換されました。生成されたDCFHは細胞膜を通過できず、細胞に簡単にロードできます。細胞内ROSは、非蛍光DCFHを緑色蛍光色のDCFに酸化しました。したがって、DCF蛍光検出は細胞内ROSのレベルを示すことができます。
両方の細胞を遊離のAtsおよびAtsをロードしたBSANPで一定期間処理すると、細胞内ROSの量も増加し、時間依存の関係を示しました。無料のAtsと比較して、SMMC-7721セルおよびAtsをロードしたBSANPで処理されたPlcセルでのROSの生成が大幅に強化されました。図4aは、AtsをロードしたBSA NPに48時間曝露したSMMC-7721細胞とPlc細胞のROSレベルが、処理したSMMC-7721細胞とPlc細胞と比較して、それぞれ1.53倍と1.28倍に増加したことを示しています。無料のAtで。これは、NPが細胞内ROSの生成を加速するという考えを支持しました。対照群および遊離Atsと比較して、AtsをロードしたBSA NPで処理した後、ER特異的色素としてのER-Tracker Blue–White DPXからの蛍光染色強度が大幅に増加し、ERストレスもトリガーされたことを示唆していますAtsをロードしたNPで処理した細胞では、対応するROSレベルの増加が見られます。この発見は、BSA NPによって媒介されるように、Atsがミトコンドリアに特異的に位置することを強調しました。これにより、細胞内の酸素フリーラジカルのレベルが大幅に上昇し、小胞体ストレスの誘導が引き起こされ、ミトコンドリア経路が活性化されて、カスパーゼ依存性の細胞アポトーシスが誘導されました。
<画像>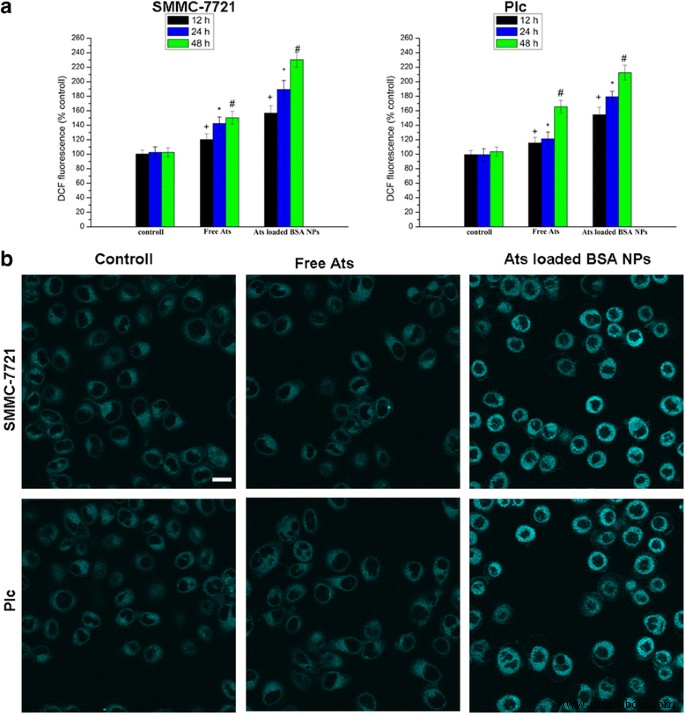
異なる時間に遊離AtsおよびAtsをロードしたBSANPで処理した細胞におけるROS生成の定量化( a )。 ER-Tracker Blue–White DPXプローブによるER染色( b )。 スケールバー 、100μm。データは平均±SD( n )として表されます =3)。 + P <0.05対12時間の対照群、* P <0.05対24時間の対照群、 # P <0.05対24時間の対照群
細胞アポトーシスと壊死の評価
細胞をアネキシンV-FITC / PI染色アッセイで処理しました。生細胞はアネキシンV-FITC / PIに結合しなかったため、蛍光は現れませんでした。アポトーシス細胞はPIに結合しませんでしたが、アネキシンV-FITCで染色され、緑色の蛍光を発しました。逆に、発癌性細胞では、細胞膜がある程度損傷し、細胞核が膨張して細かく砕け、緑色と赤色の両方の蛍光を発しました。図5aに示すように、コントロールグループと比較して、遊離のAtsおよびAtsをロードしたNPを細胞と24時間インキュベートすると、細胞内で強い緑と赤の蛍光が観察され、遊離のAtsとAtsをロードしたBSANPが示されました。誘発された腫瘍細胞の腫瘍症およびアポトーシス。特にAtsをロードしたBSANPで処理した後、アネキシンV-FITCとPIから得られた染色蛍光強度は大幅に増加し、AtsをロードしたNPで処理した細胞で発癌とアポトーシスの程度が大幅に向上したことを示唆しています。 / P> <画像>
アネキシンV-FITC / PI染色アッセイ( a )を使用して、遊離AtsおよびAtsをロードしたBSANPで処理した細胞の超微細構造変化の形態 )。 スケールバー 、100μm。それぞれ、遊離のAtsおよびAtsをロードしたBSA NPとの24時間のインキュベーション後の細胞アポトーシスおよび腫瘍症のフローサイトメーター分析( b )
初期アポトーシス(Q4)、膠質浸透圧(Q2)、壊死細胞(Q1)、および生細胞(Q3)の割合を図5bに示しました。この発見は、細胞が遊離Atsで処理されると、膠質浸透圧が24.4%と4.6%に徐々に増加し、アポトーシス率がSMMC-7721細胞とPlc細胞でそれぞれ4.9%と7.1%のままであることを示し、遊離Atsがトリガーされたことを示唆しています細胞死につながる膠質浸透圧とアポトーシスの発生。それどころか、AtsをロードしたBSA NPは、細胞のアポトーシスと腫瘍症の発生率を大幅に改善しました。アポトーシス率は、SMMC-7721細胞では10.9%、Plc細胞では11.5%に大幅に増加しました。膠質浸透圧比は、SMMC-7721細胞では29.0%に、Plc細胞では21.6%に増加しました。これは、BSA NPを介したAtsのミトコンドリア送達が、膠質浸透圧およびアポトーシス効果を増強することによって腫瘍細胞の死を加速したことを示しています。 AtsをロードしたBSANPは、アポトーシスシグナル伝達プロセスを引き起こし、ミトコンドリアを介した細胞アポトーシスのカスケード反応を促進しました。
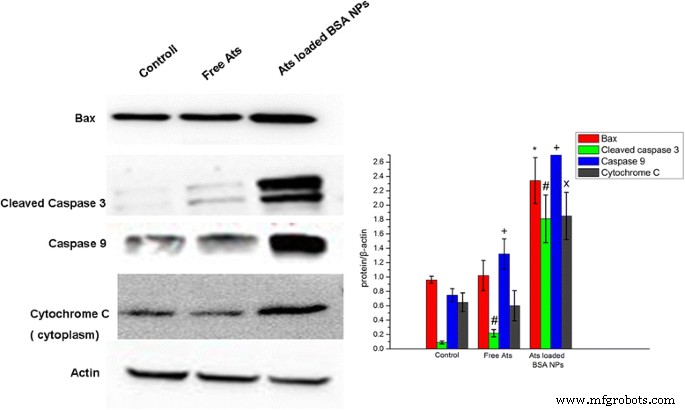
遊離のAtsおよびAtsをロードしたNPによって誘導されるアポトーシスに対する細胞死の依存性を調べるために、アポトーシスタンパク質の発現を検出するためにウエスタンブロットアッセイを実施しました。 AtsをロードしたNPで処理したSMMC-7721細胞では、Baxタンパク質の細胞内発現レベルが有意に増加していることがわかりました(図6)。この発見は、BSA NPの助けを借りて、Atsがミトコンドリアに蓄積され、ミトコンドリアの機能障害を引き起こしたことを示唆しました。細胞質のBaxモノマータンパク質はミトコンドリアの外膜に転写され、オリゴマー化を経て、ミトコンドリアの外膜にタンパク質チャネルを形成し、さらに膜透過性の増加をもたらしました。細胞質におけるシトクロムCの発現レベルも特に有意に増強され、カスパーゼ-3およびカスパーゼ-9の発現は上昇傾向を示すことがわかりました。したがって、ミトコンドリアの膜透過性が高いため、シトクロムCは細胞質に急速に放出され、細胞死シグナル伝達タンパク質(カスパーゼ)を活性化し、細胞アポトーシスのカスケード反応を促進します。対照的に、遊離Atsは、アポトーシス関連タンパク質とシトクロムCの発現に有意差がなく、遊離Atsがミトコンドリアを介した細胞アポトーシスを引き起こさず、主に腫瘍症に依存して細胞死を引き起こしたことを示唆しています。 BSA NPは、ミトコンドリアでの薬物の蓄積を促進し、ミトコンドリアを介したアポトーシス効果を活性化したため、ウエスタンブロット分析で示されているように、有意なアポトーシスを引き起こし、一次アポトーシス関連タンパク質の発現を増加させました。
SMMC-7721細胞における切断されたカスパーゼ-3、カスパーゼ-9、Bax、およびシトクロムCの発現レベルのウエスタンブロット分析。 * P <0.05対対照群のBaxタンパク質発現;
#
P <0.05対対照群の切断されたカスパーゼ-3発現;
+
P <0.05対対照群のカスパーゼ-9タンパク質発現;
x
P <0.05対対照群のシトクロムCタンパク質発現。データは平均±SD( n =3)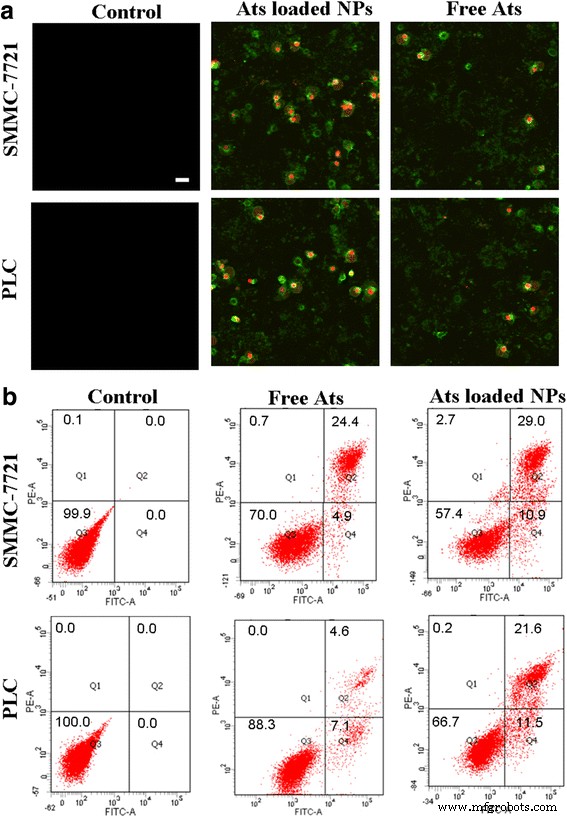
ウエスタンブロット分析
ディスカッション
腫瘍症とアポトーシスは、細胞が死ぬ2つの異なる方法を表しています。アポトーシスは、多細胞生物で発生するプログラムされた細胞死の活発なプロセスです。一方、腫瘍症は、腫れ、透過性の増加、およびしばしば壊死と呼ばれる膜破裂を特徴とするカスパーゼ非依存性細胞死を表します。この形態の細胞死は偶発的で制御されていないと考えられています。私たちの調査に基づいて、Atsが腫瘍細胞の増殖を阻害し、阻害率がAtsの用量に依存することを発見しました。 Atsは主に腫瘍の程度に依存し、細胞死を引き起こしました。また、腫瘍症の形でカスパーゼ非依存性細胞死を活性化しました。逆に、明らかな腫瘍症様死の発生とは別に、腫瘍細胞をAtsをロードしたBSA NPで処理すると、AtsをロードしたBSA NPは細胞質に内在化され、ミトコンドリア内にすばやく配置されてAtsを放出します。 NP。ミトコンドリアのAtsはROSを生成し、ERストレスを引き起こしました。さらに、ミトコンドリア膜電位を低下させ、シトクロムCを放出し、Bax、切断型カスパーゼ3、およびカスパーゼ9のタンパク質発現を促進することにより、ミトコンドリアを介したカスパーゼ依存性細胞アポトーシス経路を活性化しました。 Atsのミトコンドリア送達は、細胞死を誘発するために発癌とアポトーシスの程度を高め、それにより薬物の細胞毒性を増加させ、有意な細胞死を誘発します。
結論
簡単に説明すると、腫瘍細胞内の遊離Atは、腫瘍細胞の増殖を腫瘍細胞のような死の形で阻害するために、腫瘍の程度に強く依存していることを明らかにしました。 thus, the cytotoxicity of the drug was limited and undesirable. In contrast, Ats-loaded BSA NPs activated the mitochondrial apoptotic pathway and simultaneously triggered oncotic effects; together, they enhanced the synergistic anti-tumor efficacy of Ats. The results of this study highlighted the significance of Ats-loaded BSA NPs in the enhancement of the cytotoxic and apoptotic effects of Ats, and they further signify the role of BSA NPs in diversifying the pathways of cell death induced by Ats. Compared with free Ats, Ats-loaded BSA NPs induced greater cytotoxicity and significant cell apoptosis effects in tumor cells.
ナノマテリアル
- エミッタ接地アンプ
- 製造におけるCOVID-19の影響
- 活性酸素種の毒性を高度な腫瘍治療に調整する
- 腫瘍治療のためのドラッグデリバリーシステムとしての環境応答性金属有機フレームワーク
- Li-Nb-O化合物の調製と光触媒性能に及ぼすLi / Nb比の影響
- DNA四面体送達は、HT-29結腸癌細胞のドキソルビシン誘導アポトーシスを増強します
- メタマテリアルにおける表面プラズモンポラリトンと磁気双極子共鳴の結合効果
- 後部に黒色シリコン層を備えた結晶シリコン太陽電池の調査
- 食用菌根菌Tricholomacrassumから合成されたタンパク質でキャップされた金ナノ粒子の抗菌性、アポトーシス性、および癌細胞の遺伝子送達特性の評価
- チアカリックス[4]アレーンはミオシンATPアーゼ活性に対するZnカチオンの阻害効果を取り除く
- 農業自動化のプラスの効果