


チアカリックス[4]アレーンはミオシンATPアーゼ活性に対するZnカチオンの阻害効果を取り除く
要約
多くの女性の生殖異常は、子宮平滑筋(子宮筋層)障害によって引き起こされます。重金属は子宮平滑筋の収縮性に悪影響を及ぼします。亜鉛はほとんどの生物にとって不可欠な生体元素ですが、この元素の高用量は毒性があります。 0.5-5 mM Zn 2+ の研究 子宮からのミオシンS1ATPase活性への影響により、5 mM Zn 2+ 陽イオンは最も顕著な抑制効果を持っています。運動パラメータの計算( K m および V max 、 ATP )は、5 mM Zn 2+ の存在下でミオシンによって触媒される加水分解ATPの見かけの最大速度を明らかにしました。 1.6倍減少しました。 Кの値 m Zn 2+ の存在下でのミオシンS1によるATP加水分解の場合 減少する傾向がありますが、統計的には変化しません。子宮ミオシンS1ATPase活性は、Mg 2+ の濃度に依存しないことが確認されました。 5 mM Zn 2+ の存在下 。また、テトラヒドロキシチアカリックス[4]アレーン-テトラスルホホスホネート(C-798)およびテトラヒドロキシチアカリックス[4]アレーン-テトラホスホネート(C-800)は、5 mM Zn の存在下でミオシンS1ATPase活性をコントロールレベルに回復させることが実証されました。 2+ 。これらのチアカリックス[4]アレーン保護効果の作用を回復する最も可能性の高いメカニズムの1つは、インキュベーション培地から重金属カチオンをキレート化する能力に基づいています。 C-798およびC-800のミオシンS1領域への分子ドッキングは、これらのチアカリックス[4]アレーンがATPase活性部位近くのミオシンアミノ酸残基によってZnカチオン結合と相互作用できることを示しました。したがって、チアカリックス[4]アレーンは、このカチオンとミオシンS1の間の相互作用を弱める可能性があります。得られた結果は、高濃度の亜鉛による中毒の場合の薬理学的化合物としてこのチアカリックス[4]アレーンを使用することを目的としたさらなる研究に使用できると推測されました。
背景
重金属による環境汚染の問題と、それらが生物に与える影響を減らす方法の探求は関連しています[1、2]。
亜鉛は、ほとんどの生物にとって不可欠な生体元素です。 [3]。亜鉛イオンは、重要な代謝機能を実行する複数のタンパク質と複合体を形成します。亜鉛イオンは、50を超える異なる生化学的(生理学的)反応を触媒する少なくとも300の金属酵素の成分です[4、5] 。
ただし、亜鉛は重金属です。それは、2つの有毒金属であるカドミウムと水銀とともに、元素の周期表のグループIIbにあります。それにもかかわらず、亜鉛は人間に対して比較的無毒であると考えられています[6]。この要素は、過剰な用量でのみ有害です[7]。
経口LD 50 米国国立医学図書館のTOXNETデータベースによると、亜鉛の場合は3 g / kg体重に近いです。カドミウムの10倍以上、水銀の50倍です[6]。人間のこの微量元素の通常の濃度の過剰は、最も一般的には、それらの組成に余分な亜鉛を含む薬や生物学的に活性な添加物の摂取によって引き起こされます。それは、亜鉛でコーティングされた、または完全に亜鉛の容器に保管された食品を食べた結果としての亜鉛による中毒の個々の症例として記録されました。酸化亜鉛、塩化物、硫酸亜鉛は業界で広く使用されています ガラスの生産のため;人工繊維、亜鉛塗料、セラミック、マッチ、歯科用セメントの製造。紙パルプ産業、木材保存、錫メッキ、はんだ付けに使用されます。
高濃度の亜鉛摂取は免疫応答を変化させました[8]。いくつかの疫学研究によると、脳内のZn、Al、Cu、およびFeのレベルの上昇は、アルツハイマー病の発症または進行を促進する可能性があります[9、10]。
重金属は、胎児期の始まり、初期の発達、成熟などのさまざまな段階で女性の生殖に影響を与える可能性があります。重金属の陽イオンはまた、不妊症、不妊症、子宮内胎児発育遅延、自然流産、奇形、先天性欠損症、出生後の死亡、早期老化、学習および行動障害の原因となる可能性があります[11、12]。
子宮収縮機能は、ミオシンが酵素活性、すなわちATPを加水分解する能力を示すタンパク質複合体(アクトミオシン)の活性に関連しています。サブフラグメント-1(S1またはヘッド)の触媒ドメインに局在するミオシンATPaseは、ATPのマクロ作動性結合に蓄積された化学エネルギーを機械的運動に変換します。その結果、ミオシンはアクチンフィラメントに沿って移動し、筋収縮を引き起こします。したがって、ミオシンによって触媒されるATP加水分解は、子宮筋層機能の分子メカニズムにおける重要なプロセスの1つと見なされています[13、14]。
ミオシンサブフラグメント-1はミオシン重鎖のN末端部分であり、ATP-ase部位とアクチン結合部位を含むN末端球状モーター(触媒)ドメインと調節ドメインの2つのドメインで構成されています。またはアクチンフィラメントに沿ったミオシンの動きに関与するレバーアーム。ミオシンモータードメインのコアは、α-ヘリックスに囲まれた中央の7本鎖βシートによって形成されています。中央のβシートの7本の鎖のうち6本を占める大きな構造ドメインは、通常、上部50 kDaドメイン(U50)と呼ばれます。大きな裂け目は、465から590までのアミノ酸残基によって形成される明確に定義された構造的な下部50kDaドメイン(L50)から上部50kDaドメインを分離します。ミオシンのアクチン結合領域とヌクレオチド結合部位は上にありますヌクレオチド結合ポケットの後ろにヌクレオチドのリン酸部分がある7本鎖βシートの反対側。 Pループ、スイッチ1、およびスイッチ2は、大きな裂け目の頂点に近い上部50kDaドメインにあります。 3つのヌクレオチド結合モチーフはすべて、ヌクレオチド結合ポケットの後ろにあるヌクレオチドのリン酸部分と接触し、γ-リン酸センサーとして機能します[15]。
私たちの以前の研究では、重金属カチオンが子宮筋層の収縮特性に悪影響を与える可能性のある子宮平滑筋のミオシンATPアーゼ活性を阻害することがわかりました[16、17]。
子宮の収縮性に対する重金属の悪影響には、これらの有害な影響を排除できる薬理学的物質の開発が必要です。
カリックスアレーンは現在、さまざまな生化学的プロセスの将来の人工エフェクターとして研究者の注目を集めています。これらの化合物は、カップ型の構造を持つ合成大環状フェノールオリゴマーです。それらの上縁と下縁は、さまざまな化学置換基で機能化することができます。 Calix [4]アレーンは、4つの官能化アレーンフラグメントによって形成され、かなり柔軟な大環状化合物のコンフォメーションを特徴としています。 Calix [4] arenesは、マトリックスの毒性が低く、細胞に浸透する能力を示しました。したがって、これらの化合物は、新しい有効な薬剤を開発するための有望な薬剤と見なされています[18、19]。
このような物質の有望なクラスは、大環状分子プラットフォームに金属錯化基を有する水溶性チアカリキサレン[18]です。カリックスアレーンは、(生体)金属カチオンと超分子錯体を形成する能力があるため、重金属の抽出剤として生物医学研究でも使用されています[20、21、22]。
テトラヒドロキシ-チアカリックス[4]アレーン-テトラスルホネート(С-798)がPb 2+ の阻害効果を排除することを以前に示しました。 、Cd 2+ 、およびNi 2+ ブタ子宮筋層からのミオシンS1によって触媒されるATP加水分解について[23]。
この研究は、高濃度の亜鉛カチオンの効果と、テトラヒドロキシ-チアカリックス[4]アレーン-テトラスルホネート(С-798)およびテトラヒドロキシ-チアカリックス[4]アレーン-テトラホスホネート(C-800)とのそれらの共同作用をミオシンに及ぼすことを研究することを目的とした。子宮からのS1ATPase活性。この研究は、子宮ミオシンの酵素活性に対する高濃度の亜鉛の悪影響を排除するこれらのチアカリキサレンの能力をテストするために必要でした。
チアカリックス[4]アレーンC-798およびC-800は、それぞれ4つのアニオン性スルホン酸基と4つのホスホン酸基で上縁が修飾された4つのフェノールフラグメントによって形成されたカップで構成されています。両方のチアカリックス[4]アレーンは、下縁に高密度に位置するヒドロキシル基と二価硫黄原子を持っており、安定した金属錯体を形成して重金属をキレート化することができます[21](図1)。
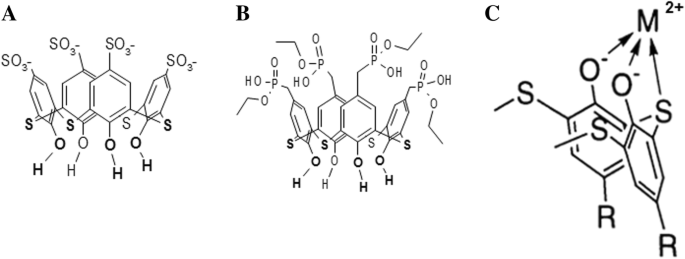
テトラヒドロキシ-チアカリックス[4]アレーン-テトラスルホネート(C-798)の化学構造( a )、テトラヒドロキシ-チアカリックス[4]アレーン-テトラホスホネート(C-800)( b )、および下縁(反転位置)に金属カチオンを有するチアカリキサレンのキレート錯体のスキーム( c )
この研究は、子宮筋層のミオシンATPaseと子宮ミオシンATPaseの阻害剤または活性化因子(エフェクター)であるカリックス[4]アレーンとの相互作用に焦点を当てた、ウクライナ国立学士院のパラディン生化学研究所と有機化学研究所の共同プロジェクトの結果です。
結果
ミオシンS1ATPase活性のZnへの依存性 2+ 濃度
子宮からのミオシンS1ATPase活性に対する最も顕著な阻害効果は、Znカチオンに対して5 mM(43±8%、M±SD)であることがわかりました。 Zn 2+ の濃度範囲 インキュベーション培地(3 mM ATP、5 mM Mg 2+ を含む)では0.5〜5mMでした 、および0.01 mM Ca 2+ )。 100%は、Znカチオンを添加しない場合のATPase活性の値です(コントロール)(図2)。したがって、ミオシンS1 ATP加水分解に対するZnカチオンの悪影響は、5mMのZn 2+ を使用してさらに研究されました。 。
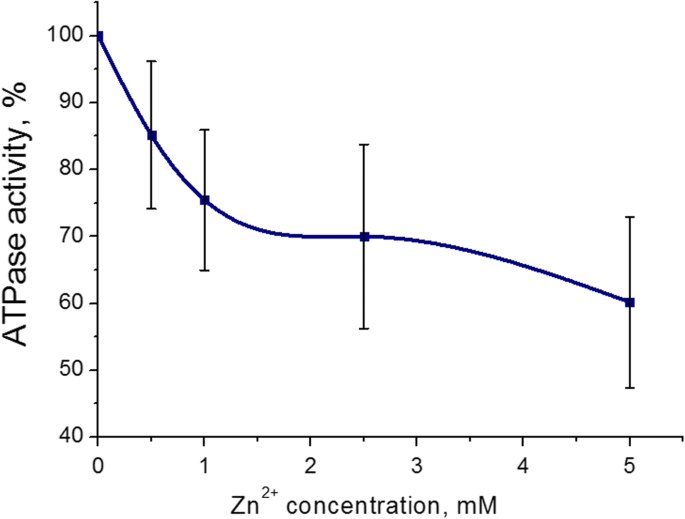
0.5〜5.0 mMの濃度のZnカチオン(M±SD、 n )の存在下での子宮筋層からのミオシンS1ATPase活性 =6)。 100%は、Znカチオンを添加しない場合のATPase活性の値です
ミオシンS1ATPase活性の存在下でのATP濃度への依存性5mM Zn 2+
Zn 2+ ミオシンS1ATPase活性の基質(ATP)への親和性への影響を調べた。固定されたMgCl 2 で、インキュベーション培地のATP濃度を0.5から5mMに上げる コントロールと5mM Zn 2+ の存在下の両方での濃度(5 mM) 3 mMATPでのATPase活性の最大値を示すドーム型のグラフが作成されました。亜鉛の存在下でのこのピークでの酵素活性の値は、コントロールの値よりも30%低かった(図3)。ミオシンS1ATPase活性の、コントロールおよび5 mM Zn 2+ の存在におけるATP濃度への依存性のグラフ 上昇部分では、Lineweaver–Burk法[27]に従って線形化されました。運動パラメータの計算、つまりミカエリスの虚数定数( K m )およびATPに対するミオシンS1 ATPaseの最大速度( V max、ATP ) V max、ATP 5 mM Zn 2+ の存在下でのミオシン酵素活性の測定 1.6倍減少しました(コントロールおよびZn 2+ の存在下でタンパク質1mgあたり38±7および22±6nmol Pi / min それぞれ、 n =5)。 Кの値 m ミオシンS1によるATP加水分解の場合、統計的に変化しませんが、減少する傾向があります(コントロールでは0.49±0.15 mM、Zn 2+ の存在下では0.38±0.12mM; M±SD; n =5)。
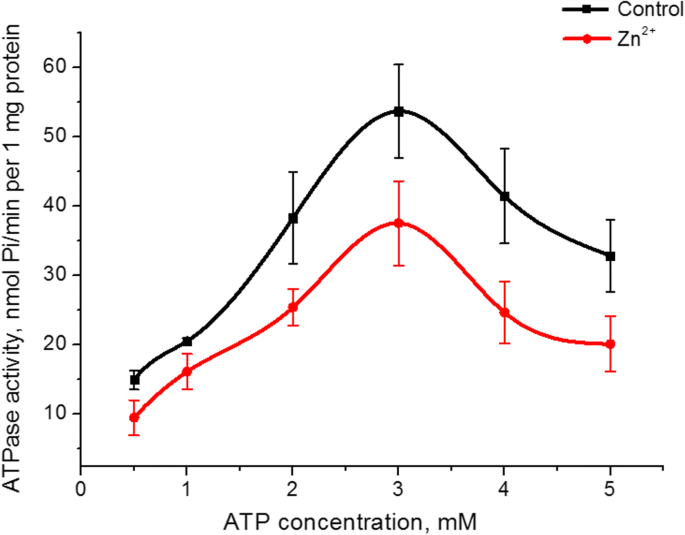
5 mM Zn 2+ の存在下での子宮からのミオシンS1ATPase活性に対する0.5〜5 mMATPの影響 コントロールとの比較(M±SD、 n =5)
ミオシンS1ATPase活性のMgへの依存性 2+ 存在下での濃度5mM Zn 2+
5 mM Zn 2+ の効果 Mg 2+ 子宮ミオシンのATPアーゼ活性への濃度依存性を研究した。 Mg 2+ を増やす 5 mM Zn 2+ の存在下での固定ATP濃度(3 mM)でのインキュベーション培地中の濃度0.5〜5 mM ミオシンS1ATPase活性の変化は生じません。同時に、Mg 2+ コントロール(標準状態)におけるATPase活性の濃度依存性が検出されました。コントロールのミオシンATP加水分解の最高レベルは、3 mM Mg 2+ で達成されました。 (図4)。したがって、子宮のミオシンS1酵素活性は、Mg 2+ の濃度に依存しません。 高濃度のZn(5 mM)の存在下で。
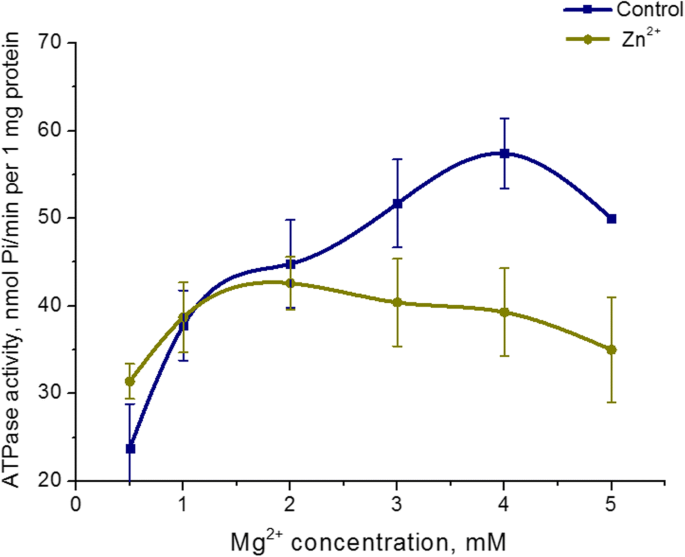
ミオシンS1ATPase活性のMg 2+ への依存性 5 mM Zn 2+ の存在下での濃度 コントロールとの比較(M±SD、 n =6)
ミオシンS1のZn結合部位
コンピューターシミュレーションは、Znカチオンがミオシンヘッドにいくつかの結合領域を持っていることを示しています。それらの1つは、上部と下部の50 kDaサブドメインの間の裂け目の下部にあり、ヌクレオチド結合部位の近くで、Pループのすぐ近くにあります。 Zn 2+ は、酸素原子Glu177(結合長0.23および0.39 nm)、Ser178酸素原子(結合長0.31 nm)、およびArg236(結合長0.32 nm)と配位しています。
他のZn結合領域は、上部50 kDaサブドメイン(Leu218-Asp463、Glu605-Phe621)の下部にあり、スイッチ1(Gly233-Phe246)とPループの近くにあります。 Zn 2+ Glu327酸素原子(結合長0.21 nm)、酸素原子Glu326(結合長0.34 nm)、および酸素原子Asp323(結合長0.32 nm)と協調できます。 Znカチオンは、スイッチ2と接触する領域のミオシンS1とも相互作用し、Glu 465(0.24 nm)、Asp468(結合長0.31 nm)、およびLeu653(結合長0.37 nm)と相互作用する場合があります。この結合領域は、アクチン結合部位と、上部と下部の50kDaサブドメインの間の裂け目にあります。この裂け目の底はATP結合ポケットにあります。これらの結合Zn 2+ ミオシンS1ドメインは、ATPの結合と加水分解に重要な役割を果たします。これらの領域は、ATP加水分解部位からアクチン結合表面にエネルギーを伝達する過程で複雑なコンフォメーション変化を起こします。
Thiacalix [4]アレーンはZn 2+ の抑制効果を排除します ミオシンATPアーゼ活性について
50 mM Tris-HClバッファー(pH 7.2)中の100マイクロモルのC-800またはC-798溶液を、5 mM Znカチオンを含むインキュベーション培地に添加して、Zn 2+ の悪影響を除去しました。 子宮平滑筋ミオシンS1のATPアーゼ活性に関する研究対照として、亜鉛および/またはチアカリックス[4]アレーンをインキュベーション培地に添加せずに酵素活性として使用した。化合物C-800は子宮筋層のミオシンS1のATPase活性に影響を与えないことが示されました(図5)。ただし、化合物C-798は、ミオシンS1 ATPase活性に対して小さな阻害効果を示します。これは、特定の量のMg 2+ の抽出に関連している可能性が最も高いです。 [21]、インキュベーション培地からの活性中心でのATP結合とその加水分解に不可欠。それにもかかわらず、100μMのC-798とC-800は、5mMのZn 2+ の抑制効果を取り除きました。 ミオシンS1によって触媒されるATP加水分解のプロセスについて。
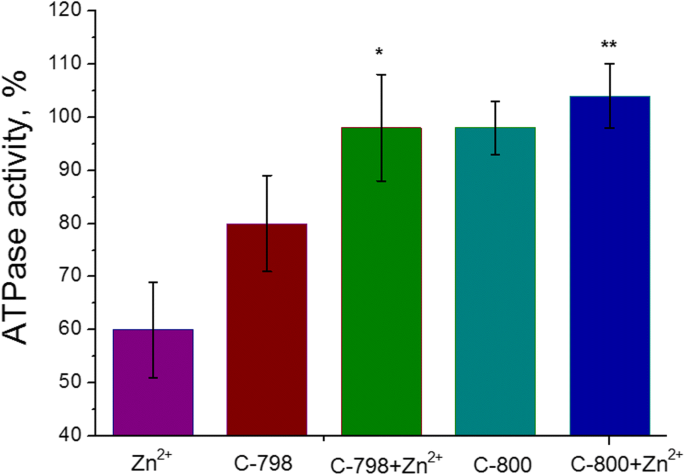
5 mM Zn 2+ の存在下でのミオシンS1ATPase活性に対する100μMC-798およびC-800の効果 (M±SD、 n =5–6)。 100%は、Znカチオンを添加しない場合のATPase活性の値です。 「Zn」と「Zn + C-798」の違い、および「Zn」と「Zn + C-800」の値の違いは、統計的に有意です( p <0.05)、それぞれ*および**として表示されます
Zn 2+ の存在下でのミオシンS1ATPase活性に対するC-798およびC-800回復作用の考えられるメカニズム
Zn 2+ のミオシンS1ATPase活性に対するC-798およびC-800回復作用の最も可能性の高いメカニズムの1つ 存在は、チアカリックス[4]アレーンがZnカチオンに結合し、その結果、これらのカチオンをインキュベーション培地から排除する能力である可能性があります。これらのチアカリックス[4]アレーンが、すでにミオシンに結合している亜鉛カチオンに結合できるかどうかは興味深いことでした。
コンピューターシミュレーションは、芳香環の間に硫黄原子を橋渡しするチアカリックス[4]アレーンC-798およびC-800が、フェノール基間の水素内結合によって安定化された「コーン」のコンフォメーションにあることを示しました。これらのカリックス[4]アレーンのエネルギー最小化構造が得られました。最小化後のC-798の総エネルギーは64.5kcal / molでした。 calix [4] areneリム(特に下部)にイオン化された基が存在すると、ホスト-ゲストの相互作用の総エネルギーに対する静電相互作用の寄与が大幅に増加します。また、C-798-Zn 2+ の「最小化」も実施しました。 繁雑;その総エネルギーは83kcal / moleでした。
C-798は、ループPの領域でタンパク質に以前に結合していたZnカチオンと協調するミオシンS1の構造に埋め込まれていました。この場合、Zn 2+ 下縁の酸素原子と相互作用し、C-798の硫黄を橋渡しします(O3、0.21 nm、S1、0.30 nm、O2、0.34 nm)。 Zn 2+ Pループのアミノ酸残基からある程度逸脱し、Glu177酸素原子(結合長0.43 nm)との相互作用を弱めます(図6)。
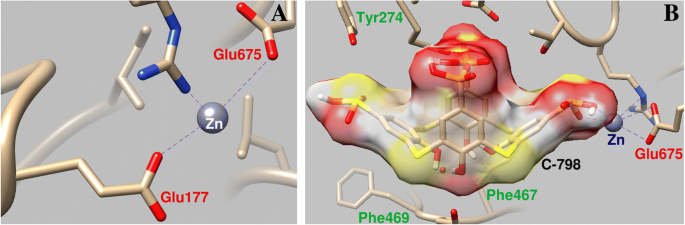
Zn 2+ の相互作用の幾何学的パラメータ ミオシンS1のPループ領域( a )およびこの領域とのZnカチオンの協力に対するC-798の影響( b )
ミオシンのATP結合領域の「空洞」でのC-798の固定は、いくつかのアミノ酸残基の関与によって起こります。特に、チアカリキサレンの疎水性バスケットは、Phe467およびPhe469のミオシン芳香族アミノ酸残基によって固定されています。チアカリキサレンの負に帯電した酸素原子は、Arg570、Asn572、およびHis689の正に帯電したアミノ酸残基と相互作用します。
Zn 2+ の変化に対するC-798の影響の研究 ミオシンATP結合部位の近くの領域でのドッキング中の位置は、Zn 2+ チアカリックス[4]アレーンの存在下では、3番目のスルホニル基(O16–0.26 nm; O15–0.27 nm)の酸素原子と相互作用し、酸素原子Asp134およびGlu326とはほとんど相互作用せず、Glu327との配位ははるかに弱くなります。 (結合長0.43 nm)(図7)。この場合、チアカリキサレンはタンパク質の「空洞」に固定されており、いくつかのアミノ酸残基が関与しています。特に、チアカリキサレンスルホニル基の負に帯電した酸素原子は、Lys188、Lys195、およびGln221のミオシンの正に帯電したアミノ酸残基と相互作用します。
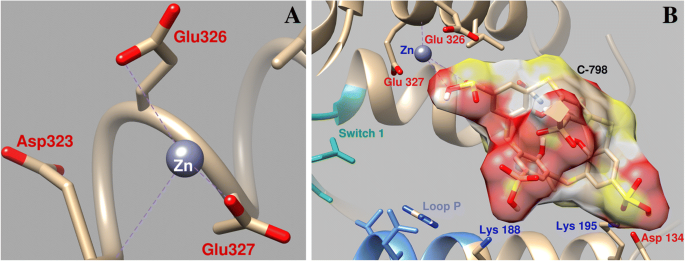
Zn 2+ の相互作用の幾何学的パラメーター ミオシンスイッチ1とミオシンS1のPループの近くの領域( a )およびZn 2+ の協力に対するC-798の影響 この領域で( b )
Znカチオンの条件付きエネルギー球は、C-798に酸素原子と硫黄原子が存在するため、静電相互作用の広がりの表面と接触し、いくらか重なり合うことが示されています。これは、研究対象の陽イオンが、上部および下部のC-798クラウンの負に帯電した原子と十分に密接に相互作用していることを示しています。チアカリキサレンはそれ自体にZn 2+ を引っ張る可能性があります 、その結果、陽イオンと酵素のアミノ酸残基との相互作用が弱まります。
その結果、Zn 2+ を含むミオシンS1領域でのC-798ドッキングの結果 結合、C-798官能基とZnカチオンとの相互作用の可能性を示します。この場合、ミオシンS1のアミノ酸残基とのZnカチオン結合は著しく弱くなり、それらの間の距離は増加します。その結果、ミオシンのATPase活性に対するZnカチオンの悪影響を排除することができます。
また、S1ミオシンのATP結合領域に近い領域にドッキングして、陽イオンZnの形状を変更する、calix [4] areneC-800の効果のコンピューターシミュレーションを実施しました。同時に、Zn 2+ Arg236(0.37 nm結合長)、Glu675(0.41 nm結合長)、および上部機能性カリックス[4]アレーン残基の原子(H26、2.36 nm、H30、2.96 nm、C30、0.31 nm、O5、 3.7 nm、O16、0.4 nm、O7、0.48 nm、およびO11、0.51 nmの結合長)。 Zn 2+ C-798と同様に、calix [4] areneC-800と相互作用します。陽イオンもこの領域の以前の結合部位から引き抜かれ、Asp468(0.2 nmの結合長)および下部カリックスアレーンクラウンの原子(C4、1.96 nm、C14、2.07 nm、S3、2.16 nm、O2)と接触しています。 、2.26 nm、C3、2.97 nm、C20、3.10 nm、O4、4.2 nm、C14、3.0 nm、O3、4.1 nm)
ミオシンサブフラグメント-1領域にドッキングするC-798およびC-800は、これらのチアカリックス[4]アレーンがATPase活性部位近くのミオシンアミノ酸残基に結合するZnカチオンと相互作用できることを示しました。したがって、それらの保護効果は、これらの陽イオンとミオシンS1の間の相互作用の弱体化の結果である可能性があります。
ディスカッション
多くの女性の生殖異常は、子宮平滑筋(子宮筋層)障害によって引き起こされます。重金属は子宮平滑筋の収縮性に悪影響を及ぼします。重金属亜鉛は、ほとんどの生物にとって不可欠な生体元素です。この元素の高用量は有毒です[5]。ミリモル濃度のZn 2+ に関するいくつかの調査 生物については以前に説明されています[32]。 5 mM Znは、子宮からのミオシンS1ATPase活性に対して最も顕著な阻害効果があることがわかりました。したがって、ミオシンS1 ATP加水分解に対するZnカチオンの悪影響は、この濃度のZn 2+ でさらに研究されています。 。 ATPに対するミオシンS1ATPaseの速度論的パラメーターの計算により、 V max、ATP 5 mM Zn 2+ の存在下でのミオシン酵素活性の測定 1.6倍減少しました。 Кの値 m ATP加水分解の場合、減少する傾向がありますが、統計的に変化しません。
生理学的条件のミオシンはMg 2+ です 依存性ATPアーゼ。マグネシウムカチオンは、ミオシン活性部位でのATP結合、およびATP加水分解に関与しています。 Mg 2+ 酵素の活性部位で、ミオシンアミノ酸残基Thr-186およびSer-237の側鎖、ならびにATP分子のβ-およびγ-リン酸基と配位し、β-およびγ-を形成します。二座複合体および活性水分子との複合体であり、そのうちの1つはγ-リン酸ATPに対して求核剤攻撃を実行します[33、34]。 Mg 2+ ATPの負に帯電したリン基と相互作用し、それらを分極化することで、末端のγ-リン酸に対する求核攻撃を促進します[14]。
ミオシンS1ATPase活性は、Mg 2+ の存在に敏感ではないことがわかりました。 5mM濃度のZn 2+ 亜鉛が培養培地に存在しなかった場合の対照とは対照的[35、36]。
ミオシンATPase活性は、金属カチオンの性質に依存し、それらのイオン半径とよく相関します。イオン半径Mg 2+ およびZn 2+ ソリューションは非常に似ています(それぞれ0.070nmと0.076nm)[37]。したがって、Zn 2+ の相互作用 Mg 2+ との陽イオン -ミオシンの結合部位が可能です。したがって、Mg 2+ -結合部位はZn 2+ で占められている可能性があります その高濃度の陽イオン。このような条件でのミオシンS1のATPase活性は、マグネシウムカチオンに鈍感である可能性があります。ミオシンには、Mg 2+ に対して2つの高親和性部位が含まれています 、およびMg 2+ これらの部位に結合すると、筋収縮中のエネルギー伝達プロセスにおいて重要な生理学的役割があります。まだいくつかのMg 2+ があります -ミオシン分子のATPase部位に加えて、マグネシウムイオンの結合エネルギーとその親和性が異なる結合部位[35]。したがって、Zn 2+ ATPの結合と加水分解に影響を与えるミオシンS1の他の機能的に重要な部位にも結合できます。
コンピューターシミュレーションは、ZnカチオンがATP結合部位の近くに位置するミオシンヘッドにいくつかの結合領域、すなわちPループと上下の50kDaサブドメインスイッチ2を持っていることを示しています。これらの結合Zn 2+ ミオシンS1ドメインは、ATPの結合と加水分解に重要な役割を果たします。これらの領域は、ATP加水分解部位からアクチン結合表面にエネルギーを伝達する過程で複雑なコンフォメーション変化を起こします。
Zn 2+ によって得られた分析結果 ミオシンS1へのドッキングは、このカチオンのミオシン分子への結合における重要な役割が、酵素アミノ酸残基の負に帯電した基、特にGluおよびAspとの相互作用を果たしていることを示しています。
毒性濃度のZn 2+ の有害な影響 ミオシンS1ATPase活性の陽イオンは、この金属の作用を排除できる薬理学的化合物の探索を必要とします。私たちの研究の対象は、安定した金属錯体を形成して遷移金属と重金属をキレート化できるテトラヒドロキシチアカリックス[4]アレーン-テトラスルホネート(C-798)とテトラヒドロキシチアカリックス[4]アレーン-テトラホスホネート(C-800)でした(図。 1)。 C-798およびC-800の上部大環状クラウンには、それぞれ4つのアニオン性スルホン酸基または4つのホスホネートが含まれており、アミノ酸フラグメントの正に帯電した窒素原子との静電接触により、チアカリキサレンの良好な水溶性とタンパク質分子への接着を提供します[21 ]。
ミオシンS1領域にドッキングするC-798およびC-800は、これらのチアカリックス[4]アレーンがATPase活性部位近くのミオシンアミノ酸残基に結合するZnカチオンと相互作用できることを示しました。したがって、それらの保護効果は、これらの陽イオンとミオシンS1の間の相互作用の弱体化の結果である可能性があります。得られた結果は、高濃度の亜鉛による中毒の場合の薬理学的化合物としてこのチアカリックス[4]アレーンを使用することを目的としたさらなる研究に使用できると推測されました。
結論
高濃度(5 mM)のZnカチオンは、子宮からのミオシンS1ATPase活性を阻害しました。 Znの抑制された影響は、5 mM Zn 2+ の存在下でミオシンS1によって触媒される加水分解ATPの最大速度の低下に関連しています。 。 Кの値 m ATPは減少する傾向がありますが、統計的に変化しません。
テトラヒドロキシチアカリックス[4]アレーン-テトラスルホホスホネート(C-798)およびテトラヒドロキシチアカリックス[4]アレーン-テトラホスホネート(C-800)は、5 mM Zn 2+ の存在下でミオシンS1ATPase活性をコントロールレベルに回復させました。 。
Znカチオンは、ATPase活性部位の近くにあるミオシンS1にいくつかの結合領域を持っています。 Zn 2+ を含むミオシンS1領域へのC-798およびC-800のドッキング 結合、これらのチアカリックス[4]アレーン官能基と結合したZnカチオンとの相互作用の可能性を示します。ミオシンS1のアミノ酸残基とのZnカチオン結合は著しく弱くなり、それらの間の距離は増加します。その結果、ミオシンのATPase活性に対するZnカチオンの悪影響を排除することができます。
得られた結果は、高濃度の亜鉛による中毒の場合に、このチアカリックス[4]アレーンを薬理学的化合物として使用することを目的としたさらなる研究に使用できると考えられます。
メソッド
試薬
次の試薬を使用しました:血清アルブミン、EGTA、EDTA、ATP、アスコルビン酸、トリス、トリシン、ジチオスレイトール、アクリルアミド、(Sigma、米国)、グリシン(Merck、ドイツ)、N、N'-メチレンビスアクリルアミド(Acros Organics、ベルギー) )N、N、N '、N'-テトラメチレンジアミン(Reanal、ハンガリー)、および国内生産の試薬(Rグレード)。溶液は、Crystal Bioシステム(Adrona、ラトビア)で精製された水で調製されました。水のコンダクタンスは0.1μS未満でした。溶液中の二価金属カチオンの濃度は、モール法によって決定されました。
アクトミオシンとミオシンサブフラグメント-1の分離
アクトミオシンは、[17]に記載されているように、修正されたBarany法によってブタの子宮平滑筋から分離されました。ミオシンS1は、ブタのアクトミオシンから修正鈴木法[24]によって得られました。サンプルの純度はPAAG-SDS電気泳動によって制御されました[25]。
ATPase活性アッセイ
ATPase activity of myosin S1 was determined in a 96-well plate at 37 °C in an incubation medium (total volume 0.1 ml) of the following composition (mM):Tris-HCl buffer (pH 7.2), 20; KCl, 100; CaCl2 , 0.01; MgCl2 , 5; and ATP, 3 (standard conditions). Protein (myosin S1) concentration was 20 μg/ml. Incubation time was 5 min. Samples containing all components of the incubation medium without myosin S1 were taken as control of non-enzyme hydrolysis of ATP. The amount of inorganic phosphate released during ATP hydrolysis reaction was determined by the Chen method [26] by the measurement of optical absorbance of the solution at 820 nm using a microplate reader μQuwant (Biotek @ Instruments, Inc., USA) and specified as Pi nmol/min per 1 mg of protein.
The Zn 2+ and thiacalix[4]arene effects on the ATPase activity of myosin S1 were studied using standard incubation medium with solutions of ZnCl2 and thiacalix[4]arenes at the corresponding concentrations. The value of ATPase activity in the absence of ZnCl2 and/or calix[4]arenes in the incubation medium was taken as 100% (control).
Kinetic and Statistical Analysis
The values of the imaginary constant of Michaelis (K m ) and maximal rate of myosin S1 ATPase for ATP (V max, ATP ) were calculated using the graph of the dependence of ATPase activity on the ATP concentration according to Lineweaver–Burk method [27]. Statistical processing of the obtained data was performed using standard methods of variation statistics. Experimental data were analyzed by using the standard software “MS Office” and “Statistica 4.5.” The statistical comparisons were performed using two-way analysis of variances (ANOVA).
Thiacalix[4]Arene Synthesis and Characterization
Tetrahydroxy-thiacalix[4]arene-tetrasulphonate and tetrahydroxy-thiacalix[4]arene-tetraphosphonate were synthesized and characterized using NMR techniques and IR spectroscopy in the Phosphoranes Chemistry Department of the Institute of Organic Chemistry, NAS of Ukraine. Infrared and NMR spectroscopy confirmed the structure of these synthesized thiacalix[4]arenes. This thiacalix[4]arenes were dissolved in water.
Computer Modeling
Computer modeling of the interaction between ligands (thiacalix[4]arenes, Zn 2+ , model bindings) and receptor (myosin S1) was performed using AutoDock software, version 4.2 [28]. We used the three-dimensional enzyme structure with the 1b7t identifier in RSCB PDB in our research [29]. Computer modeling of the thiacalix[4]arene structural peculiarities was carried out using HyperChem 7.01. Molecular dynamics calculations were performed by the MM2 method with the semi-empirical methods (CNDO).
Program AutoDockTools was used for preliminary “processing” of interacting molecules. One hundred runs of Lamarkian genetic algorithms (population size, 100; the maximal number of energy evaluations, 10 6 ) were conducted. To analyze and visualize the docking results, we used the programs Chimera [30] and Yassara [31]. Calculation of the minimal total binding energy was implemented considering Van der Waals forces, electrostatic and hydrophobic interactions, and hydrogen bonds. The optimal ligand positions in the complex “receptor-ligand” were determined according to the energy values obtained by docking software calculator for binding energy in complex “receptor-ligand.” Thus, we selected a series of complexes with the lowest total energy and then calculated the optimal geometry of the complexes and determined the most energetically preferred arrangement of the ligands in the space of myosin subfragment-1 binding domain.
略語
- C-798:
-
Tetrahydroxythiacalix[4]arene-tetrasulfosphonate
- C-800:
-
Tetrahydroxythiacalix[4]arene-tetraphosphonate
- CNDO:
-
Complete Neglect of Differential Overlap (methods)
- K m :
-
Michaelis constant, the substrate concentration at which the reaction rate of the enzyme is half of the maximal velocity
- L50:
-
Lower 50-kDa domain of myosin
- LD50 :
-
Lethal dose is the amount of an ingested substance that kills 50% of a test sample
- MM2:
-
A class of force fields
- Myosin S1:
-
Myosin subfragment-1
- NASU:
-
National Academy of Science of Ukraine
- PDB:
-
Protein Data Bank
- P-loop:
-
Phosphate-binding loop of myosin
- RCSB:
-
Research Collaboratory for Structural Bioinformatics
- U50:
-
The upper 50-kDa domain of myosin
- V max :
-
Maximal velocity of the enzyme
- V max, ATP :
-
V max for ATP
ナノマテリアル