


M1マクロファージ由来のエクソソームマイクロRNA-326は、NF-κBシグナル伝達経路を介して肝細胞癌細胞の進行を抑制します
要約
蓄積された証拠は、M1マクロファージ由来のエクソソームに由来するマイクロRNA(miR)が肝細胞癌(HCC)の進行を調節できることを示しています。ただし、HCCに対するM1マクロファージ由来のエクソソームに由来するmiR-326の効果は報告されていません。したがって、本研究の目的は、HCC細胞の進行を調節する際のM1マクロファージからのエクソソームmiR-326のメカニズムを調査することでした。 RT-qPCRは、HCC細胞株でmiR-326の発現を検出しました。 HCCにおけるmiR-326の発現はトランスフェクションによって変化し、CD206およびNF-κBの発現、細胞増殖、コロニー形成、遊走、アポトーシス、および浸潤に対するmiR-326の影響が検出されました。その後、エクソソームはM1マクロファージから分離されました。 RT-qPCRは、M1マクロファージ由来のエクソソームにおけるmiR-326の発現を同定しました。 M1マクロファージ由来のエクソソームにおけるmiR-326の発現はトランスフェクションによって変化しました。 M1マクロファージ由来のエクソソームをHCC細胞と共培養して、HCC細胞の生物学的進行に対するそれらの影響を解明しました。最後に、invitro実験を行ってinvitro結果を検証しました。 MiR-326はHCC細胞で減少し、M1マクロファージ由来のエクソソームが豊富でした。 miR-326をアップレギュレートすると、HCC細胞の増殖、コロニー形成、遊走、浸潤、CD206およびNF-κBの発現が抑制され、アポトーシスが促進され、 invivo でHCC腫瘍の増殖が抑制されます。 、miR-326をダウンレギュレートすると、逆の効果が見られました。 M1マクロファージ由来のエクソソームは、HCC細胞の増殖、コロニー形成、遊走、浸潤、CD206およびNF-κBの発現を阻害し、アポトーシスを増強しました。一方、miR-326の過剰発現は、HCC細胞に対するM1マクロファージ由来のエクソソームの効果を増強しました。 M1マクロファージ由来のエクソソームmiR-326は、NF-κBの発現をダウンレギュレートすることにより、増殖、遊走、浸潤を抑制し、HCCのアポトーシスを促進することが明らかになっています。
はじめに
肝細胞癌(HCC)は、世界で5番目に多い癌であり、最も一般的な原発性肝癌です[1]。中国国立がん登録のデータから明らかなように、原発性肝がんの死亡率は3番目に記載されており、発生率は一般的な悪性腫瘍の中で4番目です[2]。 HCCの主な危険因子は、C型肝炎ウイルスとB型肝炎ウイルスの慢性感染、アフラトキシンに汚染された食品、肥満、喫煙、大量のアルコール摂取、2型糖尿病です[3]。経動脈的化学塞栓療法は、中期段階のHCCの確立された治療法であり、中期または進行期のHCC患者の大多数の生存率を改善します[4]。現在、HCCの診断は主に血清バイオマーカーと画像技術に依存しています[5]。 HCC関連の5年生存率はわずか約60%であり、発生率は近年年々上昇しています[6]。それを考えると、正確な治療標的を研究することは、HCCの治療における優先事項です。
マクロファージは、エフェクター細胞であり、免疫系の主要な調節因子であり、組織のリモデリングと修復に多大な機能を発揮し、invivoでほとんどすべての組織で代謝機能を調整します[7]。 M1マクロファージは腫瘍促進効果を発揮し、HCC細胞の運動性を高めることが明らかになっています[8]。エクソソームは、直径40〜150nmの円板状小胞です[9]。 Xu et al。によると、エクソソームマイクロRNA(miRNA)は、標的細胞での遺伝子発現を調節することにより、HCCの増殖、浸潤性、転移、および薬剤耐性に機能します[10]。ある研究では、エクソソームを含むmiR-326が多発性硬化症の潜在的な臨床標的になる可能性があることが示されています[11]。 miRNAは、特定の配列におけるmRNAの分解と翻訳の遮断によって、タンパク質にコードされた多数の遺伝子発現を調節することにより、癌遺伝子または腫瘍阻害剤のいずれかとして機能する可能性があります[12]。ある研究では、miR-326は、細胞周期の進行を妨げるだけでなく、アポトーシスを促進することでHCC細胞の成長を抑制し、さらに上皮間葉転換の表現型を減少させることで細胞浸潤を抑制することが議論されています[13]。別の研究では、miR-326がHCC患者の治療の潜在的な治療標的になる可能性があることが報告されています[14]。したがって、この現在の研究では、HCC細胞の浸潤と遊走を調節するエクソソームmiR-326のメカニズムについて議論しました。
材料と方法
倫理声明
すべての動物実験は、国際委員会による実験動物の管理と使用に関するガイドに準拠していました。プロトコルは、吉林大学第3病院の施設内動物管理使用委員会によって承認されました。
マクロファージの誘導と同定
ヒト単球細胞株THP-1(昆明動物学研究所、CAS、昆明、中国)は、10%の熱不活化ウシ胎児血清(10%熱不活化ウシ胎児血清(Gibco、CA、USA; Thermo Fisher Scientific、MA、USA))で培養されました。 FBS)。 THP-1細胞を150ng / mLのホルボール12-ミリステート13-アセテート(PMA; P8139、Sigma-Aldrich、SF、CA、USA)と反応させ、RPMI培地で24時間インキュベートして、M0マクロファージを得ました。次に、誘導前後の細胞形態をライト染色で観察した。 THP-1細胞と誘導されたマクロファージを5μLのPBSに再懸濁し、スライドガラスに滴下し、ライトの色素溶液で染色し、緩衝液と1:2で混合しました。 10分間染色し、流水ですすいだ後、細胞を顕微鏡で観察した。さらに、M0マクロファージマーカー(CD68およびCD206)は、逆転写定量的ポリメラーゼ連鎖反応(RT-qPCR)によって測定されました。次に、20 ng / mLインターフェロン(IFN)-γ(#285-IF; R&D Systems、MN、USA)および10 pg / mL LPS(#8630; Sigma-Aldrich)との18日間のインキュベーションにより、マクロファージをM1マクロファージに誘導しました。 h。 M1マクロファージマーカー(IDO1およびIL-12 p35)は、RT-qPCRによって調べられました[15]。
エクソソームの抽出
エクソソームは、エクソソーム分離キット(ExoEasy Maxi Kit、Qiagen、ヒルデン、ドイツ)によって抽出されました。マクロファージの上清を無菌条件下で15mLの遠心分離管に集め、0.8μMのろ過フィルムでろ過しました。各グループの細胞上清にXBPバッファー(1:1)を添加し、exoEasyメンブレンアフィニティー遠心分離カラムで500 g で遠心分離しました。 。細胞に10mLのXWPバッファーを補充し、3000〜5000 g で遠心分離しました。 。 exoEasyメンブレンアフィニティー遠心分離カラムを400μLのXE溶出バッファーでハッチングし、500 g で遠心分離しました。 。溶出バッファーをexoEasyメンブレンアフィニティー遠心分離カラムに移し、500 g で遠心分離しました。 。溶出バッファーは4°Cで24時間保存し、同定に使用しました。残りは−80°Cで保管されました。
TEM観察およびナノ粒子追跡分析(NTA)
上記で得られたエクソソームを炭素支持膜銅メッシュに滴下し、2%リンタングステン酸を加えた。サンプルをTEMで観察し、最適な画像を収集して分析しました。
PBS中の不純物と粒子を0.22μMミクロポーラスフィルターで除去しました。エクソソームの粒子密度に応じて、ろ過されたPBSを適切な濃度に希釈し、Nanosight NS300ナノ粒子検出器(Malvern、Westborough、MA、USA)を使用して検出しました。
同定後、miR-326阻害剤およびmiR-326阻害剤ネガティブコントロール(NC)、miR-326ミミックおよびmiR-326ミミックNCでトランスフェクトされたマクロファージ由来のエクソソームをエクソソーム分離キット(Invitrogen)で抽出しました。
RT-qPCR
全RNAはTrizolReagent(Thermo Fisher)によって抽出され、リアルタイムPCRはSYBR-Green PCR Master Mix(Roche)とABI 7500 Real-Time PCR System(Life Technologies、Grand Island、NY、USA)を使用して実行されました。 )。プライマー配列を表1に示します。定量分析は2 -△△Ct の方法を使用して実施しました。 。
ウエスタンブロット分析
細胞とエクソソームの総タンパク質を抽出しました。タンパク質濃度は、ビシンコニン酸(BCA)キット(Boster Biological Technology Co. Ltd.、Wuhan、Hubei、China)によって決定されました。タンパク質にサンプルバッファーを添加し、95°Cで煮沸し、各ウェルに30 µgをロードしました。タンパク質を10%ポリアクリルアミドゲル電気泳動(Boster Biological Technology)で分離し、ポリフッ化ビニリデンメンブレンにエレクトロブロッティングした後、5%ウシ血清アルブミン(BSA)でシールしました。膜は、CD63(1:1000、Developmental Studies Hybridoma Bank、アイオワ大学、アイオワ州、アイオワ州、米国)、CD181(1:1000、R&D Systems)、GAPDH(1:2000、Jackson ImmunoResearch Laboratories、米国アイオワ州)および西洋ワサビペルオキシダーゼで標識された二次抗体(1:500、Jackson ImmunoResearch Laboratories)を使用。画像はOdysseyデュアルカラー赤外線蛍光スキャンイメージングシステムによって取得され、バンドのグレー値はQuantityOne画像分析ソフトウェアによって測定されました。
細胞培養とスクリーニング
正常肝細胞株HL-7702およびヒトHCC細胞株BEL-7404、HepG2、SMMC-7721およびQGY-7703を選択し、10%ウシ胎児血清(FBS)、ペニシリン(100 U / mL)を含むGibco RPMI Media1640で培養しました。 )およびストレプトマイシン(100mg / mL)。 MiR-326の発現はRT-qPCRによって検出され、適切な細胞株がスクリーニングされました。
エクソソームのラベル付けとエクソソームの取り込み
エクソソームを250μLの希釈液Cで再懸濁し、エクソソームの膜に損傷を与えないように穏やかに粉砕しました。 PKH67色素(1μL、Sigma-Aldrich)を250μLの希釈液Cに添加して500μLに到達させ、インキュベートしました。溶液に500μLの1%BSAを添加し、37°Cで1分間インキュベートしました。エクソソームは、120,000 g での遠心分離によって得られました。 4°Cで2時間。 PKH67で標識されたエキソソームは、120,000 g での遠心分離によって得られました。 4°Cで2時間。エクソソームは、光を避けて6 mLRPMI-1640培地で再懸濁しました。次に、標識されたエクソソームをHCC細胞と12時間共培養しました。その後、培地を除去し、PBSで3回、5分/時間洗浄し、HCC細胞に内部吸収されなかった蛍光標識エクソソームを完全に洗い流した。エクソソームを4%パラホルムアルデヒドで固定し、4'-6-ジアミジノ-2-フェニルインドールで染色しました。密封後、レーザー共焦点顕微鏡で蛍光分布を観察しました。
セルのグループ化と処理
HepG2細胞とSMMC-7721細胞を12ウェルプレートに0.5–1×10 6 で播種しました。 細胞/ウェル。 50〜60%のコンフルエンスで、細胞にLipofectamine 2000(Invitrogen、カリフォルニア州カールズバッド)をトランスフェクトしました。 HepG2細胞は、miR-326-mimicグループ(miR-326 mimicでトランスフェクト)とNC-mimicグループ(miR-326 mimic NCでトランスフェクト)に分配されました。 SMMC-7721細胞は、miR-326阻害剤グループ(miR-326阻害剤でトランスフェクト)とNC阻害剤グループ(miR-326阻害剤NCでトランスフェクト)に割り当てられました。 miR-326-mimic、miR-326-inhibitorおよびそれらのNCは、トランスフェクションのためにLipofectamine2000と混合されました。何も処理していないHepG2細胞とSMMC-7721細胞をブランクグループとして設定しました。 miR-326-mimic、miR-326-inhibitorおよびそれらのNCは、Guangzhou RibBio Co.、Ltd。(中国、広州)によって考案および構成されました(表1)。
<図>M1マクロファージ由来のエクソソームとHCC細胞の共培養
M1マクロファージ由来のエクソソーム懸濁液のタンパク質濃度をBCA法で検出し、50μgのタンパク質を含む対応するエクソソーム懸濁液の量を計算しました。 HepG2細胞とSMMC-7721細胞を12ウェルプレートに1×10 5 で播種しました。 ウェルあたりの細胞数/ mL。 HepG2細胞は4つのグループに分けられました:コントロールグループ(エキソソームと共培養されていないHepG2細胞)、エキソソーム(Exo)グループ(M1マクロファージ由来のエキソソームと共培養されたHepG2細胞)、Exo-miR-326模倣グループ(HepG2 miR-326ミミックでトランスフェクトされたM1マクロファージ由来エキソソームと共培養された細胞)、Exo-NC-ミミックグループ(miR-326ミミックNCでトランスフェクトされたM1マクロファージ由来エキソソームと共培養されたHepG2細胞)。 SMMC-7721細胞も4つのグループに割り当てられました:ブランクグループ(エキソソームと共培養されていないSMMC-7721細胞)、Exoグループ(M1マクロファージ由来のエキソソームと共培養されたSMMC-7721細胞)、Exo-miR-326-阻害剤グループ(miR-326阻害剤をトランスフェクトしたM1マクロファージ由来エキソソームと共培養したSMMC-7721細胞)、Exo-NC阻害剤グループ(miR-をトランスフェクトしたM1マクロファージ由来エキソソームと共培養したSMMC-7721細胞) 326阻害剤NC)。
3-(4,5-ジメチルチアゾール-2-イル)-2,5-ジフェニルテトラゾリウムブロミド(MTT)アッセイ
細胞をトリプシンで剥離し、細胞密度が4×10 4 の96ウェルプレートに播種しました。 ウェルあたりの細胞。それぞれ12、24、36、48、60時間培養した後、培地を廃棄した。 500μLの0.5g / L MTT溶液とインキュベートし、細胞に200μLのジメチルスルホキシド溶液を添加し、粉砕して孵化しました。光学密度(OD、490 nm)値は、マイクロプレートリーダーで測定しました。
コロニー形成アッセイ
24時間培養し、トリプシンで剥離し、細胞を35 mmの小さな皿に播種し、1皿あたり300個の細胞を入れました。溶液は3日ごとに交換されました。 10日間の培養後、細胞を40g / L -1 で固定しました。 パラホルムアルデヒドおよび1g / L -1 で染色 クリスタルバイオレット溶液と乾燥。コロニー数(50個以上の細胞)は顕微鏡下で計算されました。
トランスウェルアッセイ
セル(1×10 5 )200μLのブランク培地で懸濁した。実験は、トランスウェルチャンバー(コーニンググラスワークス、コーニング、ニューヨーク州、米国)の指示に従って実施された(マトリゲルは侵入実験には必要であったが、移動実験には必要ではなかった)。 RPIM 1640(10%FBS、600μL)を下部チャンバーに追加しました。上部チャンバーと下部チャンバーは、マトリゲル(BD Biosciences、フランクリンレイクス、ニュージャージー州、米国)でプレコートされたトランスウェルメンブレンによって分離されました。 24時間培養し、チャンバーを95%アルコールで固定しました。クリスタルバイオレット溶液で染色した後、顕微鏡下で5つの視野で細胞を観察しました。
フローサイトメトリー
細胞周期:細胞はトリプシンによって剥離されました。セル(1×10 6 )0.5 mL PBSで懸濁し、粉砕して単一懸濁液にしました。氷上で4.5mLの予冷した70%エタノールと混合し、細胞を3000 g で遠心分離しました。 、5 mL PBSでリンスし、3000 g で再度遠心分離します。 。続いて、0.2mgのRnaseAを含む1mLのPI / Triton X-100染色液(20μgPI/ 0.1%Triton X-100)で細胞を懸濁しました。細胞周期はフローサイトメトリーによって検出されました。
細胞アポトーシス:トリプシン処理された細胞(1×10 6 )1 mL PBSで懸濁し、粉砕し、3000 g で遠心分離しました。 。細胞をインキュベーションバッファー(10 mmol / L Hepes / NaOH、pH 7.4、140 mmol / L NaCl、5 mmol / L CaCl 2 )で順番にすすいだ。 )そして3000 g で遠心分離します 。次に、細胞を100μLのマーキング溶液でインキュベートし(FITC-アネキシンVおよびPIをインキュベーションバッファーに加えて1μg/ mLに到達)、3000 g で遠心分離しました。 、インキュベーションバッファーで1回洗浄し、蛍光(SA-FLOUS)溶液で孵化しました。細胞アポトーシスはフローサイトメトリーによって検出されました。フローサイトメトリーの波長は488nmで、FITC蛍光は515 nmのバンドフィルターで検出されましたが、PIは560nmを超えています。結果はコンピューターによって自動的に分析されました。
ヌードマウスの腫瘍異種移植
4〜6週齢の40匹のマウス(吉林大学、吉林省、中国)をランダムに8つのグループに分け、各グループに5匹のマウスを配置しました。マウスは特定病原体除去グレードの動物実験室で1週間飼育され、飼料、クッション、ウォーターボトルは時間内に交換されました。マウスの健康状態を毎日観察する必要があります。 1週間後、HCC細胞を細胞懸濁液に調製し、0.1 mLの細胞懸濁液(1×10 6 )で首と背中に皮下注射しました。 )。腫瘍の成長は3〜5日後に観察されました。ヌードマウスの体重を4日ごとに測定し、腫瘍体積を測定しました。ヌードマウスは注射の20日後に安楽死させた。
統計分析
すべてのデータは、SPSS 17.0ソフトウェア(SPSS Statistics、シカゴ、イリノイ、米国)によって解釈されました。測定データは平均±標準偏差として示されました。 2つのグループ間の比較は、 t によって作成されました。 -テスト、複数のグループ間の比較は一元配置分散分析(ANOVA)によって評価されました。 P 値<0.05は統計的に有意な差を示しました。
結果
M1マクロファージとエクソソームの識別
ライト染色を利用して、PMAによって誘導されたTHP-1細胞の形態を観察した。誘導前のTHP-1細胞の量は少なく、齲蝕原形質の割合が高かったことが描かれました。誘導後の細胞の形態は不規則であり、体積が大きくなり、齲蝕原形質の割合が減少した。細胞質はより豊富で、水色で、豊富な粒子といくつかの液胞がありました。核は紫がかった赤で、しばしば片側に傾いており、細胞がマクロファージの典型的な形態学的特徴を持っていることを示しています(図1a)。
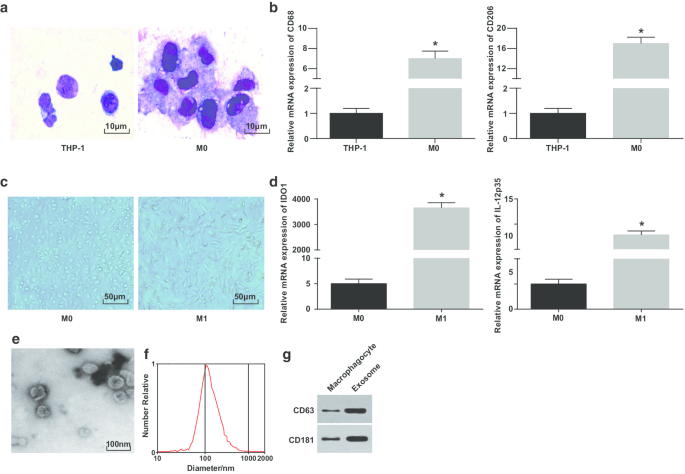
M1マクロファージとエクソソームの同定。 a PMA処理前後のTHP-1細胞の形態を観察するためのライト染色。 b RT-qPCRによって検出されたTHP-1細胞およびPMA処理THP-1細胞におけるCD68およびCD206の発現。 c マクロファージとM1マクロファージの形態。 d RT-qPCRによって検出されたM0マクロファージおよびLPSおよびINF-γ処理M0マクロファージにおけるIDO1およびIL-12p35の発現。 e エクソソーム観察のためのTEM。 f NTAによるエクソソームの粒度分布の検出。 g CD63およびCD181のタンパク質バンド。パネルbでは、* P <0.05対THP-1細胞;パネルdでは、* P <0.05対M0マクロファージ。測定データは、平均±標準偏差( N )として表されました。 =3)、2つのグループ間の比較はt検定によって行われました
マクロファージの誘導が成功したことをさらに検証するために、誘導前後のCD68およびCD206の発現をRT-qPCRでテストしました。 CD68およびCD206の発現はPMA誘導以降上昇していることが明らかになり、PMAがTHP-1細胞をM0マクロファージに正常に誘導したことを示しています(図1b)。次に、LPSとINF-γの誘導により、M0マクロファージがM1マクロファージに極性化されました。形態、およびマクロファージのM1型マクロファージ表面マーカーIDO1およびIL-12 p35の発現を観察し、テストしました。 M0マクロファージは、円形、楕円形、または紡錘形を示す、さまざまな不規則な付着形態を示しました。 IFN-γによって刺激されたマクロファージの形態は、より多くの仮足と突起および紡錘形で示されました(図1c)。 RT-qPCRは、LPSおよびINF-γ処理後、M1マクロファージがマーカー(IDO1およびIL-12p35)の増加を示したことを概説しました(図1d)。
続いて、マクロファージに由来するエクソソームがTEMによって観察された。マクロファージ由来のエクソソームは豊富で、形状は円形または楕円形で、膜状の構造、均一なサイズ、汚染物質が少ないことが明らかになりました(図1e)。 NTAは、MODE曲線の中央にピークがあり、線形が滑らかなエクソソームは、直径がより集中しており、不純物が少ないことを示しました(図1f)。ウエスタンブロット分析は、マクロファージと比較して、特定のマーカータンパク質CD63およびCD181の発現がマクロファージに由来するエクソソームで上昇したことを報告しました(図1g)。これらの結果は、単球をマクロファージに分化させ、M1マクロファージに極性化することに成功したことを示しています。
M1マクロファージ由来のエクソソームはmiR-326をHCC細胞に送達し、HCC細胞でのmiR-326の発現に影響を与えます
M1マクロファージに由来するエクソソームがmiR-326をHCC細胞に輸送したかどうかを検証するために、HepG2およびSMMC-7721細胞をエクソソームと共培養しました。蛍光顕微鏡下での4時間のトランスフェクションで、HepG2およびSMMC-7721細胞によって多数のエクソソームが同化されたことが認識できました(図2a、b)。
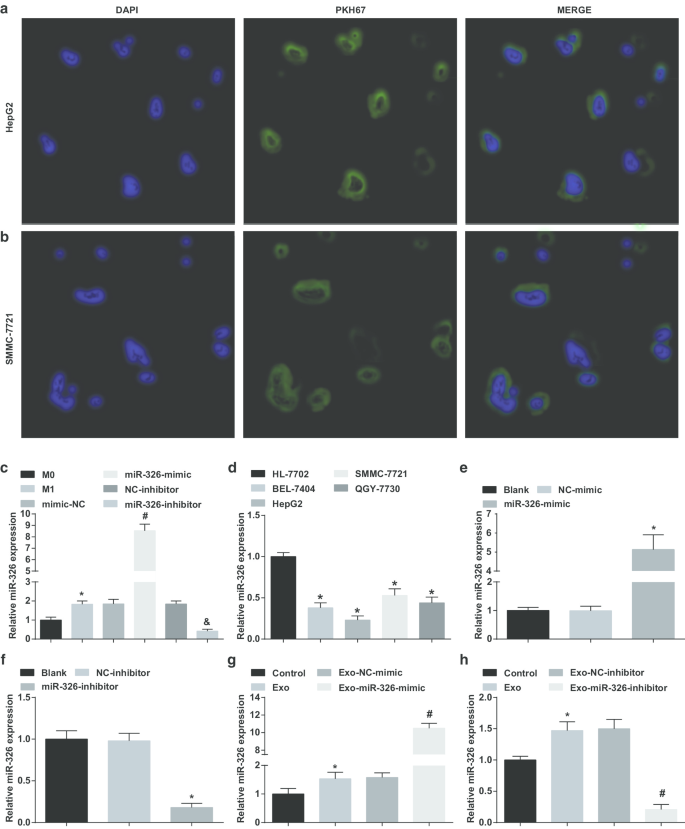
M1マクロファージ由来のエクソソームはmiR-326をHCC細胞に送達し、HCC細胞でのmiR-326の発現に影響を与えます。 a HepG2細胞によるM1マクロファージ由来のエクソソームの取り込み。 b SMMC-7721細胞によるM1マクロファージ由来のエクソソームの取り込み。 c INF-γおよびLPS誘導前後の各グループにおけるマクロファージエクソソームのmiR-326発現の比較。 d HCC細胞株(BEL-7404、HepG2、SMMC-7721、QGY-7703)およびRT-qPCRによって検出されたヒト正常肝細胞HL-7702細胞株におけるmiR-326の発現。 e RT-qPCRは、HepG2細胞におけるmiR-326発現に対するmiR-326ミミックの効果を検出しました。 f RT-qPCRは、SMMC-7721細胞におけるmiR-326発現に対するmiR-326阻害剤の効果を検出しました。 g RT-qPCRは、HepG2細胞におけるmiR-326の発現に対するmiR-326ミミックをトランスフェクトしたM1マクロファージ由来のエクソソームの影響を検出しました。 h RT-qPCRは、SMMC-7721細胞におけるmiR-326発現に対するmiR-326阻害剤をトランスフェクトしたM1マクロファージ由来のエクソソームの効果を検出しました。パネルcでは、* P <0.05対M0マクロファージ;パネルdでは、* P <0.05対HL-7702セル;パネルeでは、* P <0.05対NC模倣グループ;パネルfでは、* P <0.05対NC阻害剤グループ;パネルgでは、* P <0.05対対照群、# P <0.05対Exo-NC-mimicグループ;パネルhでは、* P <0.05対対照群、# P <0.05対Exo-NC阻害剤グループ。測定データは、平均±標準偏差( N )として表されました。 =3)、複数のグループ間の比較は、一元配置分散分析によって行われました
続いて、miR-326-mimic / inhibitorをマクロファージにトランスフェクトし、トランスフェクションの前後にエクソソームでのmiR-326の発現を確認しました。 M0マクロファージからのエクソソームと比較して、miR-326の発現はM1マクロファージ由来のエクソソームで上昇したことが示された。上昇したmiR-326発現は、miR-326模倣治療を受けたM1マクロファージ由来のエクソソームで見られました。 MiR-326阻害剤処理により、M1マクロファージ由来のエクソソームでMiR-326の発現が低下しました(図2c)。
次に、HCC細胞株におけるmiR-326の発現をテストしました。明らかなように、miR-326の発現は、HL-7702細胞と比較してBEL-7404、HepG2、SMMC-7721、およびQGY-7703細胞で減少し、HepG2細胞は最も低い発現を示し、SMMC-7721細胞は最も高い発現を示しました。 (図2d)。
続いて、miR-326-mimicおよびmiR-326-inhibitorをそれぞれHepG2およびSSMC-7721細胞にトランスフェクトし、miR-326発現に対するそれらの影響を調べました。 MiR-326はHepG2細胞でのmiR-326発現の上昇を模倣し、miR-326阻害剤はSSMC-7721細胞でのmiR-326発現を低下させました(図2e、f)。
次に、miR-326-mimicおよびmiR-326-inhibitorでトランスフェクトされたM1マクロファージからのエクソソームを、それぞれHepG2およびSSMC-7721細胞と共培養しました。エクソソームとの共培養はHCC細胞でmiR-326発現を増加させ、miR-326-mimicでトランスフェクトされたM1マクロファージ由来のエクソソームはHepG2細胞でmiR-326発現をさらに上昇させ、miR-326阻害剤でトランスフェクトされたM1マクロファージはさらに上昇したことが強調されました由来のエクソソームは、SSMC-7721細胞でmiR-326の発現を低下させました(図2g、h)。 M1マクロファージ由来のエクソソームがmiR-326をHCC細胞に送達し、HCC細胞でのmiR-326の発現に影響を与えることが示唆されています。
M1マクロファージ由来のエクソソームmiR-326は、HCC細胞の細胞増殖とコロニー形成能力を低下させます
エクソソームmiR-326がHCC細胞の増殖に及ぼす影響を調べる際に、HCC細胞の増殖を調べるためにMTTおよびコロニー形成アッセイを実施しました。 HepG2細胞では、miR-326の回復により、細胞増殖とコロニー形成能が損なわれることが示唆されました(図3a、e、f)。 M1マクロファージ由来のエクソソームはHepG2細胞の増殖を阻害し、コロニーを形成しました。 MiR-326ミミックでトランスフェクトされたM1マクロファージ由来のエクソソームは、細胞増殖とコロニー形成能力をさらに損ないました(図3c、i、j)。
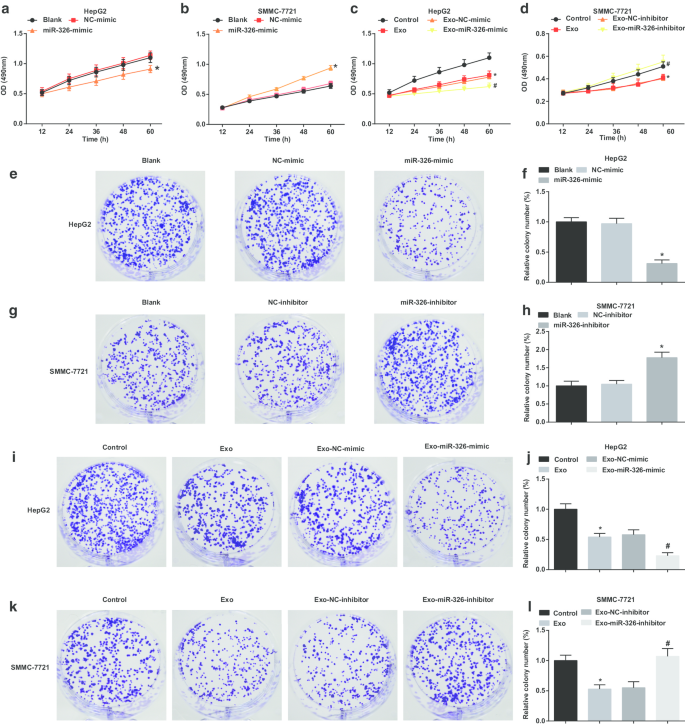
M1マクロファージのエクソソームmiR-326は、HCC細胞の細胞増殖とコロニー形成を阻害します。 a MTTアッセイは、HepG2細胞の増殖に対するmiR-326ミミックのトランスフェクションの効果を検出しました。 b MTTアッセイは、SMMC-7721細胞の増殖に対するmiR-326阻害剤のトランスフェクションの効果を検出しました。 c MTTアッセイは、HepG2細胞の増殖に対するmiR-326ミミックをトランスフェクトしたM1マクロファージ由来のエクソソームとの共培養の効果を検出しました。 d MTTアッセイは、SMMC-7721細胞の増殖に対するmiR-326阻害剤をトランスフェクトしたM1マクロファージ由来のエクソソームとの共培養の効果を検出しました。 e コロニー形成アッセイは、HepG2細胞のコロニー形成能力に対するmiR-326ミミックのトランスフェクションの効果を検出しました。 f HepG2細胞のコロニー数。 g コロニー形成アッセイは、SMMC-7721細胞のコロニー形成能力に対するmiR-326阻害剤のトランスフェクションの効果を検出しました。 h SMMC-7721細胞のコロニー数。 i コロニー形成アッセイは、HepG2細胞のコロニー形成能力に対するmiR-326ミミックでトランスフェクトされたM1マクロファージ由来のエクソソームとの共培養の効果を検出しました。 j エクソソームで処理されたHepG2細胞のコロニー数。 k コロニー形成アッセイは、SMMC-7721細胞のコロニー形成能力に対するmiR-326阻害剤でトランスフェクトされたM1マクロファージ由来のエクソソームの共培養の効果を検出しました。 l エクソソームで処理されたSMMC-7721細胞のコロニー数。パネルaとfでは、* P <0.05対NC模倣グループ;パネルbおよびhでは、* P <0.05対NC阻害剤グループ;パネルcおよびjでは、* P <0.05対対照群、# P <0.05対Exo-NC-mimicグループ;パネルdとlでは、* P <0.05対対照群、# P <0.05対Exo-NC阻害剤グループ。測定データは、平均±標準偏差( N )として表されました。 =3)、複数のグループ間の比較は、一元配置分散分析によって行われました
SMMC-7721細胞では、miR-326ノックダウンにより細胞増殖とコロニー形成能が増強されました(図3b、g、h)。 M1マクロファージ由来のエクソソームで処理されたSMMC-7721細胞では、細胞増殖とコロニー形成能が低下しました。 MiR-326阻害剤でトランスフェクトされたM1マクロファージ由来のエクソソームは、細胞増殖とコロニー形成能力をさらに促進しました(図3d、k、l)。 M1マクロファージ由来のエクソソームに由来するmiR-326がHCC細胞の増殖を妨げることが示唆されています。
M1マクロファージ由来のエクソソームmiR-326は、HCC細胞の移動と浸潤を抑制します
次に、HCC細胞の浸潤および遊走に対するエクソソームmiR-326の効果を調べた。 HepG2細胞では、miR-326の回復により、浸潤と遊走が制限されることが実証されました(図4a–c)。 M1マクロファージ由来のエクソソームはHepG2細胞の侵入と移動を妨害しました。 HepG2細胞をmiR-326-mimicでトランスフェクトされたM1マクロファージ由来のエクソソームと共培養すると、浸潤と遊走がさらに低下しました(図4g–i)。
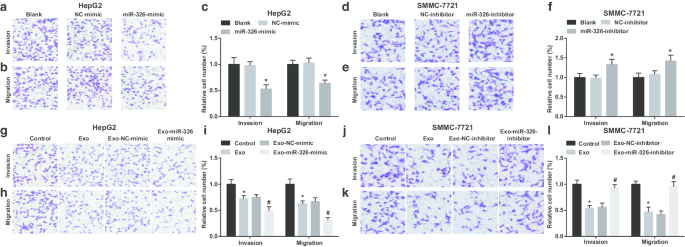
M1マクロファージのエクソソームmiR-326は、HCC細胞の遊走と浸潤を阻害します。 a トランスウェルアッセイは、HepG2細胞の浸潤に対するmiR-326ミミックのトランスフェクションの効果を検出しました。 b トランスウェルアッセイは、miR-326のトランスフェクションの効果がHepG2細胞の移動を模倣していることを検出しました。 c HepG2細胞の侵入と移動の数。 d トランスウェルアッセイは、SMMC-7721細胞の浸潤に対するmiR-326阻害剤のトランスフェクションの効果を検出しました。 e トランスウェルアッセイは、SMMC-7721細胞の遊走に対するmiR-326阻害剤のトランスフェクションの効果を検出しました。 f SMMC-7721細胞の侵入と移動の数。 g トランスウェルアッセイは、HepG2細胞の浸潤に対するmiR-326ミミックでトランスフェクトされたM1マクロファージ由来のエクソソームとの共培養の効果を検出しました。 h トランスウェルアッセイは、HepG2細胞の遊走に対するmiR-326ミミックでトランスフェクトされたM1マクロファージ由来のエクソソームとの共培養の効果を検出しました。 i M1マクロファージ由来のエクソソームと共培養されたHepG2細胞の浸潤と遊走の数。 j トランスウェルアッセイは、SMMC-7721細胞の浸潤に対するmiR-326阻害剤をトランスフェクトしたM1マクロファージ由来のエクソソームとの共培養の効果を検出しました。 k トランスウェルアッセイは、SMMC-7721細胞の遊走に対するmiR-326阻害剤をトランスフェクトしたM1マクロファージ由来のエクソソームとの共培養の効果を検出しました。 l M1マクロファージ由来のエクソソームと共培養されたSMMC-7721細胞の浸潤と遊走の数。パネルcでは、* P <0.05対NC模倣グループ;パネルfでは、* P <0.05対NC阻害剤グループ;パネルiでは、* P <0.05対対照群、# P <0.05対Exo-NC-mimicグループ;パネルlでは、* P <0.05対対照群、# P <0.05対Exo-NC阻害剤グループ。測定データは、平均±標準偏差( N )として表されました。 =3)、複数のグループ間の比較は、一元配置分散分析によって行われました
MiR-326ノックダウンにより、SMMC-7721細胞の浸潤と遊走が増強されました(図4d–f)。 M1マクロファージ由来のエクソソームで処理した場合、SMMC-7721細胞は浸潤と遊走が減少して示されました。 However, SMMC-7721 cell invasion and migration were boosted upon co-culture with miR-326-inhibitor-transfected M1 macrophages-derived exosomes (Fig. 4j–l). It is implied that miR-326 derived from M1 macrophage exosomes impedes the invasion and migration of HCC cells.
M1 Macrophage-Derived Exosomal miR-326 Promotes Apoptosis of HCC Cells
When examining the effect of exosomal miR-326 on the cell cycle and apoptosis of HCC cells, PI single staining and Annexin V-FITC/PI double staining were applied. It was illustrated that miR-326 overexpression increased cells arrested at G0/G1 phase, reduced cells arrested at S and G2/M phases and raised apoptosis in HepG2 cells (Fig. 5a–d). Co-culturing with M1 macrophages-derived exosomes increased cells arrested at G0/G1 phase, reduced cells arrested at S and G2/M phases and raised cell apoptosis of HepG2 cells. Co-cultivation with exosomes from M1 macrophages transfected with miR-326-mimic further enhanced these effects (Fig. 5i–l).

M1 macrophage exosomal miR-326 promotes apoptosis of HCC cells. a 、 b Flow cytometry detected the effect of transfection of miR-326 mimic on HepG2 cell cycle; c 、 d flow cytometry detected the effect of transfection of miR-326 mimic on HepG2 cell apoptosis; e 、 f flow cytometry detected the effect of transfection of miR-326 inhibitor on SMMC-7721 cell cycle; g 、 h flow cytometry detected the effect of transfection of miR-326 inhibitor on SMMC-7721 cell apoptosis; i 、 j flow cytometry detected the effects of co-culture of with miR-326 mimic-transfected M1 macrophage-derived exosomes on HepG2 cell cycle; k 、 l flow cytometry detected the effect of co-culture with miR-326 mimic-transfected M1 macrophage-derived exosomes on HepG2 cell apoptosis; m 、 n flow cytometry detected the effect of co-culture with miR-326 inhibitor-transfected M1 macrophage-derived exosomes on SMMC-7721 cell cycle; o , p , flow cytometry detected the effect of co-culture with miR-326 inhibitor-transfected M1 macrophage-derived exosomes on SMMC-7721 cell apoptosis. In panel b and d, *P <0.05 versus the NC-mimic group; In panel f and h, *P <0.05 versus the NC-inhibitor group; In panel j and l, *P <0.05 versus the control group, #P <0.05 versus the Exo-NC-mimic group; In panel n and p, *P <0.05 versus the controlgroup, #P <0.05 versus the Exo-NC-inhibitor group. Measurement data were depicted as mean ± standard deviation (N = 3), comparisons among multiple groups were conducted by one-way analysis of variance
In SMMC-7721 cells, miR-326 down-regulation reduced cells arrested at G0/G1 phase, elevated cells arrested at S and G2/M phases, and declined cell apoptosis (Fig. 5e–h). Untransfected M1 macrophages-derived exosomes increased cells arrested at G0/G1 phase, reduced cells arrested at S and G2/M phases, and heightened cell apoptosis. MiR-326-inhibitor-transfected M1 macrophages-derived exosomes degraded cells arrested at G0/G1 phase, elevated cells arrested at S and G2/M phases, and decreased cell apoptosis (Fig. 5m–p). Briefly, it is summarized that miR-326 derived from M1 macrophage exosomes arrests cell cycle in G0/G1 phase and induces cell apoptosis in HCC.
M1 Macrophage-Derived Exosomal miR-326 Declines CD206 and NF-κB Expression in HCC Cells
Next, the potential mechanism of miR-326 derived from M1 macrophage exosomes in the biological progress of HCC cells was explored. NF-κB is the key link between inflammation and cancer. Many regulatory proteins and miRNAs could inhibit the excessively activated NF-κB signaling to suppress cancer [16]. Such a beneficial effect may include the polarization of M2 macrophages into M1 macrophages. CD206 and NF-κB expression in HepG2 and SMMC-7721 cells was tested by RT-qPCR. It was suggested that miR-326 restoration decreased CD206 and NF-κB expression in HepG2 cells, while miR-326 knockdown enhanced CD206 and NF-κB expression in SMMC-7721 cells (Fig. 6a, b). Moreover, co-culture with M1 macrophage exosomes significantly reduced CD206 and NF-κB expression in HepG2 cells, while co-culture with M1 macrophage exosomes-overexpressing miR-326 further decreased CD206 and NF-κB expression. Treated with untransfected M1 macrophages-derived exosomes, CD206 and NF-κB expression was decreased in SMMC-7721 cells. Co-cultured with miR-326-inhibitor-transfected M1 macrophages-derived exosomes, SMMC-7721 cells were featured by heightened CD206 and NF-κB expression (Fig. 6c, d). It was concluded that miR-326 from M1 macrophage exosomes played a tumor suppressor by inhibiting NF-κB in HCC cells.

M1 macrophage exosomal miR-326 declines CD206 and NF-κB expression in HCC cells. a RT-qPCR detected the effect of transfection of miR-326 mimic on the expression of CD206 and NF-κB in HepG2 cells; b RT-qPCR detected the effect of transfection of miR-326 inhibitor on the expression of CD206 and NF-κB in SMMC-7721 cells. c RT-qPCR detected the effect of co-culture with miR-326 mimic-transfected M1 macrophage-derived exosomes on the expression of CD206 and NF-κB in HepG2 cells; d RT-qPCR detected the effect of co-culture with miR-326 inhibitor-transfected M1 macrophage-derived exosomes on the expression of CD206 and NF-κB in SMMC-7721 cells. In panel a, *P <0.05 versus the NC-mimic group; In panel b, *P <0.05 versus the NC-inhibitor group; In panel c, *P <0.05 versus the control group, #P <0.05 versus the Exo-NC-mimic group; In panel d, *P <0.05 versus the control group, #P <0.05 versus the Exo-NC-inhibitor group. Measurement data were depicted as mean ± standard deviation (N = 3), comparisons among multiple groups were conducted by one-way analysis of variance
miR-326 from M1 Macrophage Exosomes Inhibits HCC Tumor Growth In Vivo
Finally, the in vivo results were validated through tumor xenografts. As displayed, miR-326 overexpression decreased volume and weight of tumors in HepG2 cells (Fig. 7a–c). In mice transplanted with HepG2 cells co-cultured with exosomes, the treatment with M1 macrophage exosomes significantly reduced the tumor volume and weight of HepG2 cells, while treatment with M1 macrophages-overexpressing miR-326 further reduced the tumor volume and weight of HepG2 cells (Fig. 7g–i).

M1 macrophage exosomal miR-326 reduces the volume and weight of HCC tumor in vivo. a – c The effect of transfection of miR-326 mimic on the tumor volume and tumor of nude mice xenografted with HepG2 cells. d – f The effect of transfection of miR-326 inhibitor on the tumor volume and tumors of nude mice xenografted with SMMC-7721 cells. g – i The effect of co-culture with miR-326 mimic-transfected M1 macrophage-derived exosomes on the tumor volume and tumor of nude mice xenografted with HepG2 cells. j – l The effect of co-culture with miR-326 inhibitor-transfected M1 macrophage-derived exosomes on the tumor volume and tumor of nude mice xenografted with SMMC-7721 cells. In panel b and c, *P <0.05 versus the NC-mimic group; In panel e and f, *P <0.05 versus the NC-inhibitor group; In panel h and i, *P <0.05 versus the control group, #P <0.05 versus the Exo-NC-mimic group; In panel k and l, *P <0.05 versus the control group, #P <0.05 versus the Exo-NC-inhibitor group. Measurement data were depicted as mean ± standard deviation (n = 5), comparisons among multiple groups were conducted by one-way analysis of variance
In SMMC-7721 cells, miR-326 suppression increased volume and weight of tumors (Fig. 7d–f). Untransfected M1 macrophages-derived exosomes obstructed tumor growth in volume and weight of SMMC-7721 cells. Co-cultured with miR-326 inhibitor-transfected M1 macrophages-derived exosomes, SMMC-7721 cells were injected into mice and caused elevations in tumor volume and weight (Fig. 7j–l). Summarily, miR-326 derived from M1 macrophage exosomes depressed tumor growth of HCC in vivo.
ディスカッション
HCC is a common cancer that is characterized with high morbidity and mortality, difficult early diagnosis and treatment, poor prognosis and 5-year survival rate [17]. Recently, a study has highlighted that the lowly expressed lncRNA cox-2 declines the ability of M1 macrophages to suppress HCC cell growth, invasiveness, angiogenesis migration and promote apoptosis [18]. Hu et al. have discussed that miR-326 is obviously degraded in HCC tissues and cell lines, while down-regulated miR-326 is connected to the TNM stage, lymph node metastasis and differentiation of HCC patients [14]. It is customarily considered that HCC cells-derived exosomes can form a fertile environment to facilitate HCC cells growth, invasiveness and metastasis as well as development of drug resistance [19]. The current study was designed to explore the mechanism of exosomal miR-326 in regulating invasion and migration of HCC cells.
Our results indicated that miR-326 expression decreased in HCC cells but increased in exosomes. A recent study has pointed out that miR-326 expression is declined in HCC tissues [20]. Another study has presented miR-326 expression is notably reduced in HCC cell lines and tissues and its down-regulation predicts a poor prognosis in HCC [13]. It is reported that miR-326 acts a tumor-suppression role and is greatly depressed in HCC cells [21]. All these aforementioned evidences are in line with our findings. A study has purported that in comparison with the controls, miR-326 expression is raised in Tconv-derived exosomes which is observed in relapsing–remitting multiple sclerosis patients [11].
Other results emerge from our data that highly expressed exosomal miR-326 reduced cell proliferation, colony formation, migration and invasion as well as facilitated apoptosis of HCC cells in vitro and reduced the volume and weight of HCC tumor in vivo. It has been suggested previously that HCC cell growth can be suppressed via overexpression of miR-326, and HCC cell migration and invasion ability are markedly attenuated through elevating miR-326 [21]. It is reported that the up-regulated miR-326 expression suppresses HCC cell growth and invasiveness as well as stimulates cell apoptosis in vitro [14]. Besides that, a prior study has verified that overexpression of miR-326 declined tumor growth in vivo [13]. A study has revealed that ectopic expressed miR-326 markedly attenuates cell growth, and suppresses cellular migration and invasiveness in non-small cell lung cancer cell lines [22]. Moreover, it is found that miR-326 decreases profibrotic genes like MMP-9, implying its repressive function in cancer cell proliferation [23]. Also, it is presented that miR-326 represses Bcl-2 protein expression and elevates Bax expression so as to affect the apoptosis [24]. Similar to our findings, there are some miRNAs interacting with exosomes to play a role in HCC development. It is displayed that highly expressed exosomal miR-638 can repress the proliferation of HCC cells, involving the potential impact on carcinogenesis [25]. Another study also proves that HCC cells-derived exosomal miR-451a suppresses tumor angiogenesis via disrupting endothelial functions as apoptosis, tube formation, migration and permeability [26]. A prior research generally confirms that when treated with the overexpression of miR-744 exosomes, the proliferation of HCC cells is dramatically suppressed [27].
結論
To briefly conclude, our study provides evidence that M1 macrophage-derived exosomal miR-326 suppresses proliferation, migration and invasion as well as advances apoptosis of HCC cells, supplying a new insight in a novel target therapy for HCC. Due to the limited sample size and limited known researches, the exact mechanism of miR-326 is not fully elucidated, and therefore, further large-scale studies are required to illustrate the underlying mechanism.
略語
- HCC:
-
肝細胞がん
- miRNAs:
-
マイクロRNA
- FBS:
-
ウシ胎児血清
- M/CSF:
-
Macrophage colony-stimulating factor
- PBS:
-
リン酸緩衝生理食塩水
- DMEM:
-
Dulbecco’s modified eagle medium
- PPAR:
-
Peroxisome proliferator-activated receptor
- TEM:
-
透過型電子顕微鏡
- NTA:
-
Nanoparticle tracking analysis
- NC:
-
ネガティブコントロール
- RT-qPCR:
-
逆転写定量的ポリメラーゼ連鎖反応
- BCA:
-
ビシンコニン酸
- BSA:
-
ウシ血清アルブミン
- Exo:
-
Exosomes
- ANOVA:
-
一元配置分散分析
ナノマテリアル
- 誘電体メタ表面を介した帯域幅の拡大による完全なテラヘルツ分極制御
- HT29およびSPEV細胞株に対するAuナノ粒子の影響のinvitro研究
- MAPbI3ペロブスカイト太陽電池構造における斜め角度スパッタITO電極の効果
- エレクトロスピニング法による直接メタノール燃料電池用の新しい複合Tio2カーボンナノファイバー陽極触媒担体の製造と特性化
- PEG化リポソームを介したブファリンの改善された抗腫瘍効果および薬物動態
- ヒドロキシル基を介したカーボンナノチューブのスライドロール運動モードの調整
- 金ナノ粒子で阻害されたチトクロームP4503A4活性と、ヒト肝細胞癌細胞株C3Aにおけるその細胞毒性の根底にある分子メカニズムの評価
- 小さな希土類フッ化物ナノ粒子は、電気極性相互作用を介して腫瘍細胞の成長を活性化します
- 過酸化亜鉛ベースのプログラム可能な金属化セルのスイッチング故障メカニズム
- 食道扁平上皮癌に対する有効性を高めるためのオートファジー阻害剤(LY294002)と5-フルオロウラシル(5-FU)の組み合わせベースのナノリポソーム
- 胃壁細胞および腸杯細胞分泌:中国のハーブを介して下痢で増強された新しい細胞媒介invivo金属ナノ粒子代謝経路