


海洋植物プランクトンからのエキソポリマー物質放出に対する操作されたナノ粒子の影響
要約
現代のナノテクノロジーの製品であるエンジニアードナノ粒子(ENP)は、海洋環境に影響を与え、海洋生態系に深刻な脅威をもたらす可能性があります。ただし、ENPに対する海洋植物プランクトンの細胞応答はまだ十分に確立されていません。ここでは、4つの異なる珪藻種( Odontella mobiliensis )を調査します。 、 Skeletonema grethae 、 Phaeodactylum tricornutum 、タラシオシラシュードナナ )と1つの緑藻( Dunaliella tertiolecta )モデルENP処理での細胞外高分子物質(EPS)の放出:25 nm二酸化チタン(TiO 2 )、10〜20 nmの二酸化ケイ素(SiO 2 )、および15〜30 nmの二酸化セリウム(CeO 2 )。 SiO 2 が見つかりました ENPは、これらの藻類からのEPS放出を大幅に刺激することができます(200〜800%)が、TiO 2 ENP曝露は最低の放出を誘発しました。さらに、細胞内Ca 2+ の増加 濃度はENPによって引き起こされる可能性があり、EPS放出プロセスがCa 2+ を介して媒介されることを示唆しています。 信号経路。 ENPによって誘発されるEPS放出を介した細胞メカニズムをよりよく理解することで、海洋生態系への悪影響を軽減するための潜在的な予防および安全対策を開発できます。
背景
サイズが1〜100 nm(少なくとも1次元)の範囲にある人工ナノ粒子(ENP)は、プリンターのインクや塗料、洗剤、殺菌剤、コーティング、化粧品、日焼け止めローションなど、さまざまな消費財の製造に使用されています。タイヤ、コンピューターの構築、および薬物送達。 ENPの有望な適用を考えると、米国だけでNational Nanotechnology Initiative(NNI)への資金提供は2017年に14億ドルに近づきました[1,2,3]。ナノスケールでの基礎知識の確立は、第一段階でのナノテクノロジー研究コミュニティの主な焦点でした。 2009年の時点で、この新しい知識は世界市場の約4分の1兆ドルを支えており、そのうち約910億ドルはナノスケールのコンポーネントを組み込んだ米国製品に含まれていました[4]。ナノテクノロジーの急速な発展に伴い、ENPが最終的に水生システムへの道を見つけることは避けられません。環境における潜在的な毒性(例えば、活性酸素種、ROSを生成する可能性)に関するENPの主な懸念は、それらの大きくて独特な表面反応性に関連しています。しかし、天然水の複雑な環境的および生物学的要因とさまざまなENPのために、海洋生態系への実際の影響はほとんど不明のままです[1、5、6]。以前の研究では、ENPが藻類に基づく海洋生態系に重大な害を及ぼす可能性があることが示されています[7、8]。海洋生物(特に植物プランクトン)はENPと相互作用し、負の影響をもたらすことが示されています[9、10、11]。多様な分野でナノテクノロジーの利用が増加する可能性があるため、ますます多くのENPが水生環境に入る可能性があるため、ENPに対する海洋植物プランクトンの細胞応答にはさらに注意が必要です[12、13、14、15、16、17、18、19、20、 21]。
ほとんどの海洋微生物は、自家栄養か従属栄養かにかかわらず、一般に、成長阻害剤、成長促進剤、毒素、金属スカベンジャーとして、または従属栄養サイクル[22、23、24、25、26]。植物プランクトンや海洋のバクテリアから放出されるEPSは、多糖類が豊富なアニオン性コロイド状バイオポリマーであり、海洋ゲル、マリンスノー、バイオフィルムの形成、コロイドや微量元素の除去、さまざまな環境脅威からの保護に不可欠です。 ENPを含む[7、15、19、20、25、27]。さらに、EPSの分泌は、植物プランクトンがさまざまなストレスを経験したときの自然な反応であると考えられています[8]。
Ca 2+ は、多数の細胞内シグナル伝達経路に関与する一般的なセカンドメッセンジャーです。 Ca 2+ 珪藻 Amphora coffeaeformis の走化性、運動性、接着に必要です [28]。強化された細胞内遊離Ca 2+ レベルは、多くの細胞内シグナル伝達経路に関与するプロテインキナーゼCの活性化につながることが知られています[29]。 EPSの放出は珪藻の運動性と付着性に密接に関連しているため、Ca 2+ 媒介分泌プロセスは、珪藻からのEPSの放出を制御し[30]、Ca 2+ を証明する直接的な証拠 シグナル伝達、エキソサイトーシス、および相関するCa 2+ エキソサイトーシスによるシグナル伝達は、以前の研究で報告されています[31]。過去の研究では、ENPとの相互作用が細胞内Ca 2+ を変化させる可能性があることも示されています 細胞シグナル伝達に不可欠な経路[29、32、33、34]。特定の細胞内Ca 2+ 濃度の変化は、細胞のシグナル伝達および分泌プロセスにおいて重要です。ただし、二酸化チタン(TiO 2 )の報告はありません。 )、二酸化ケイ素(SiO 2 )、または二酸化セリウム(CeO 2 )細胞内Ca 2+ を変更する 植物プランクトンのレベル。
2013年、Quigg etal。 [8]は、藻類に対するENPの直接的および間接的な毒性作用を要約した。以前の実験では、ENPがEPSの集約を促進することが示されました[35]。この点で、EPSは水生生物に対する直接的なENP誘発毒性を悪化または減少させる可能性があります[7、15、36]。ただし、ENPsストレス下の植物プランクトンからのEPS放出の直接測定は報告されていません。この研究の目的は、4つの異なる珪藻種( Odontella mobiliensis )からのEPSの放出を研究することです。 、 Skeletonema grethae 、 Phaeodactylum tricornutum 、タラシオシラシュードナナ )と1つの緑藻( Dunaliella tertiolecta )ENP処理中。植物プランクトンにおけるENP誘発EPS放出の根本的なメカニズムを理解することにより、予防および安全対策の実施は、海洋生物に対する潜在的に有害な影響を軽減することができます。
結果と考察
ENPの特性評価
動的レーザー散乱(DLS)を使用して、純水に懸濁した次のENPのサイズメトリックを特徴付けました。TiO 2 、SiO 2 、およびCeO 2 。 TiO 2 では、粒子サイズの分布は7〜66nmの範囲でした。 、SiO 2 で9〜66 nm 、およびCeO 2 で12〜70 nm 。いくつかのより大きなサイズは、凝集または凝集が原因である可能性がありますが、TiO 2 の主なサイズは は25nm、SiO 2 は10〜20 nm、CeO 2 は15〜30 nmであり、メーカーの情報と一致しています(図1)。
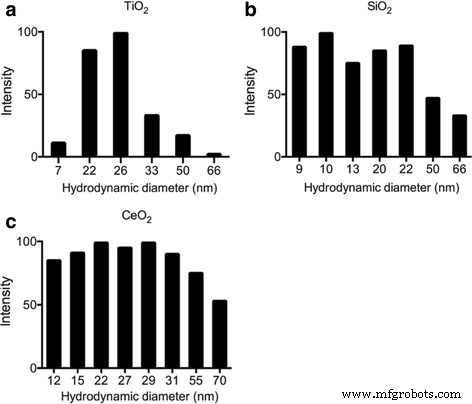
a のDLS評価によるENPの特性評価 TiO 2 、 b SiO 2 、および c CeO 2 超音波処理後のL1培地で、サイズ分布を示しています。 DLSサンプルのENP最終濃度は1μg/ mlで、測定時間は超音波処理直後の3分です
ENPは細胞内Ca 2+ を誘発します 植物プランクトンの濃度
ENPが細胞内Ca 2+ の増加を誘発できるかどうかを調査する 濃度、植物プランクトン細胞(OD 600 =0.8)にFluo-4AM色素をロードし、1 mg / mlの25nm TiO 2 に曝露しました。 、10〜20 nm SiO 2 、および15〜30 nm CeO 2 それぞれENP。細胞内Ca 2+ の変化 植物プランクトン細胞内の蛍光強度で表される濃度を150秒間モニターしました。図2a–eは、3つのそれぞれのENPの1 mg / mlがCa 2+ を増加させたことを示しています。 SiO 2 の濃度 約50〜300%、TiO 2 約40%、CeO 2 対照条件(L1培地)は変更されていませんが、約150〜200%増加しました。結果は、ENPが有意な細胞内Ca 2+ を誘発できることを示しています 植物プランクトンの反応と植物プランクトンがCa 2+ を介して異なるENPに反応することを示唆している シグナル伝達経路。私たちのデータは、細胞内Ca 2+ のわずかな変化のみを示しています TiO 2 のときのレベル 存在し、TiO 2 による実質的な植物プランクトン細胞死に起因する可能性があります 誘発毒性[37、38]。以前の調査では、TiO 2 細胞内Ca 2+ の増加を促した 濃度[34]と有意な細胞アポトーシス[39]。ただし、SiO 2 驚くべきことに、最も明白な細胞内Ca 2+ を示しました すべての植物プランクトン種で増加し、CeO 2 中間の細胞内Ca 2+ のみをトリガーできます 濃度が上がる。以前の調査では、CeOが高い 2 の可能性が示唆されていました 細胞内酸化ストレスと細胞内Ca 2+ の上昇を誘発する濃度(> 50 mg / ml) 効果は小さかったものの、レベルは、私たちの発見を支持しました[40]。また、人工海水中の各ENPのゼータ電位を測定して、表面電荷によって引き起こされる可能性のある影響に対処しました。ただし、値は低かった。測定により、ENPはほぼ中立と見なされることが示されました[41](追加ファイル1:補足データ)。これは、異種のENPが細胞内Ca 2+ を誘発することがわかった最初の報告として役立ちました。 特定の植物プランクトンの濃度が変化し、最終的には将来の研究のための新しい道が開かれます。
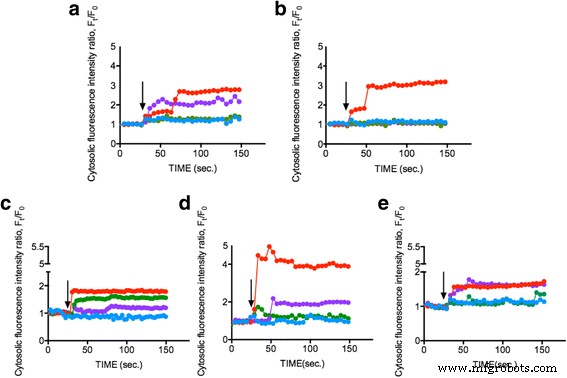
細胞内Ca 2+ の測定 異なるENPによる刺激後の濃度。さまざまな植物プランクトン細胞 a Dunaliella tertiolecta 、 b タラシオシラシュードナナ 、 c Skeletonema grathae 、 d Phaeodactylum tricornutum 、および e Odontella mobiliensis TiO 2 で処理されました 25 nm(緑)、SiO 2 10〜20 nm(赤)、CeO 2 15〜30 nm(紫)、濃度1 mg / mlおよびコントロール(青)。黒い矢印は、EPNが適用された時点(30秒)を示しています。測定値は、平均20個の個別のセルからの代表的なデータを示しています
ENPによる植物プランクトンのEPSリリース
酵素結合レクチンアッセイ(ELLA)を使用して、TiO 2 で刺激した場合の植物プランクトン細胞からのEPS放出量を評価しました。 、SiO 2 、およびCeO 2 ENP、濃度範囲はTiO 2 の以前の研究に基づいて1μg/ mlから5mg / mlです。 [42、43]およびCeO 2 [44,45,46]。 EPS分泌は、比較のために等しいベースを持つために、植物プランクトンの総DNA量(追加ファイル1:補足データ)に正規化されました。コントロールと比較すると、10〜20nmのSiO 2 Dunaliella でEPSリリースを最大550%増加させることができます 、 Thalassiosira で500% 、 Skeletonema で1000% 、 Odontella で400% 、および Phaeodactylum で900% (図3)。植物プランクトン種がTiO 2 にさらされたとき 、 Skeletonema のみであるため、EPS分泌に強い影響はありませんでした。 および Phaeodactylum 大きな変化を示しました。したがって、EPS放出データは細胞内Ca 2+ と一致しています。 濃度の結果。 TiO 2 細胞内Ca +2 と同様に、EPSの生成に大きな影響はありませんでした。 濃度は、TiO 2 の毒性のために非常に限界的な変化を示しました 植物プランクトンに。 ROSの生成と残留は、植物プランクトンのアポトーシスなどの多くの合併症を引き起こす可能性があります[47、48、49]。 CeO 2 治療、結果は Dunaliella でわずかな効果を示しました 、スケルトネマ 、 Odontella 、および Phaeodactylum 。ただし、SiO 2 Thalassiosira pseudonana で最も有意なEPS誘導を示した (約600%)および Skeletonemagrethae (約1000〜1500%)。これらのデータは、さまざまなENPが植物プランクトンおよび細胞内Ca 2+ からの特定のEPS放出を誘発する可能性があることを示しています。 変更はEPSリリースの結果とも一致します。細胞内Ca 2+ の変化を評価することによって 濃度が高い場合、Ca 2+ に直接的な関係があることは明らかです。 ENPが植物プランクトンからのEPS分泌を誘発する細胞経路。ここでの観察は、 Phaeocystis に基づく以前の研究と一致しています。 EPSリリース[31]。この結果は、植物プランクトンがCa 2+ によって調節されるさまざまなEPS放出に応答するENPを検出および区別できるという直接的な証拠を提供します。 細胞経路。
ELLAを使用することで、植物プランクトンとENPの相互作用を介してEPSの放出を決定することができました。私たちの結果は、植物プランクトンがSiO 2 と相互作用するにつれて、EPS分泌が大幅に増加したことを示しています。 Dunaliella tertiolecta の場合 、タラシオシラシュードナナ 、および Skeletonema grethae 。これらの珪藻は、SiO 2 を認識するように準備されているようです。 粒子。ただし、 Phaeodactylum tricornutum 、強いEPS分泌は見られなかった。この違いは、植物プランクトンの種とENP濃度に依存するENPによって引き起こされるEPS放出を表しています(図3)。以前の研究では、油流出は、油流出の悪影響を打ち消すために提案された大量の海洋微生物EPS放出を引き起こしました[50]。さらに、BoglaienkoとTanselは、SiO 2 粒子は油の骨材を効率的に除去することができました[51]。私たちの発見は、低毒性のSiO 2 粒子は特定の植物プランクトンからのEPS放出を誘発し、EPSの凝集を促進することで油流出の除去を促進する可能性があります。二酸化セリウムが植物プランクトンに基づく海洋生態系を乱すことが報告されたことはありません。ここでの結果は、CeO 2 を示しています ENPは、 Thalassiosirapseudonanaを除くすべての植物プランクトンに影響を与える可能性があります。 CeO 2 ENPは、SiO 2、のように、 油を軽減する用途のために、特定の植物プランクトンからのEPS放出を促進する能力があります。
結論
ENPと海洋環境の相互作用は、ナノマテリアルの現在および将来の排出によりますます重要になっています。ここでは、植物プランクトンに対するENPの主要な効果の1つとして強化されたEPS分泌を示します。また、Ca 2+ を調節することにより、さまざまな植物プランクトンがさまざまなENPストレスに対してさまざまに反応する可能性があるという証拠も提供します。 経路。ただし、海洋生態系に対するENPの完全な評価には、ナノマテリアルと海洋生物の間の相互作用の詳細な知識と理解を提供するために、さらに調査する必要があります。
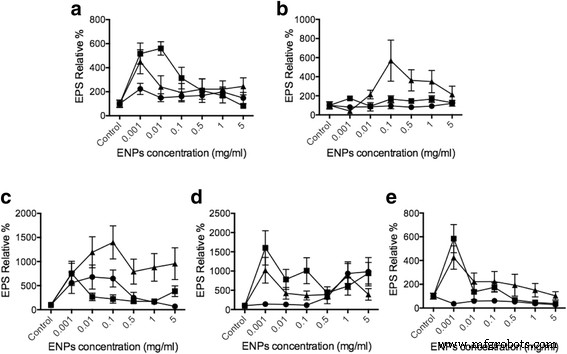
さまざまなENPによってトリガーされるEPSリリース。さまざまな植物プランクトン細胞 a Dunaliella tertiolecta 、 b タラシオシラシュードナナ 、 c Skeletonema grathae 、 d Phaeodactylum tricornutum 、および e Odontella mobiliensis TiO 2 で処理されました (円)、SiO 2 (三角形)、CeO 2 (四角)、それぞれ5 mg / mlおよび1mg / ml、0.5 mg / ml、0.1 mg / ml、10μg/ ml、1μg/ mlの濃度( n =3)
メソッド
植物プランクトンの文化
Odontella mobiliensis のバッチ培養 (CCMP597)、 Dunaliella tertiolecta (UTEX999)、 Skeletonema grethae (CCMP775)、 Phaeodactylum tricornutum (UTEX646)、タラシオシラシュードナナ (Provasoli-Guillard海洋植物プランクトン培養コレクション、West Boothbay Harbour、MN、USA)は、L1海洋培地(Sigma、MO、USA)で、100μmolm -2の14:10(明:暗)サイクルで培養されました。 s -1 無菌条件下で24°C。培養物の増殖期は、血球計算盤で細胞を数えることによって決定されました。
ナノ粒子と特性評価
すべてのENP、TiO 2 、SiO 2 、CeO 2 (Sigma-Aldrich、MO、USA)は、使用前に純水中で超音波処理しました。 ENPは、テスト前にフィルター処理されたL1媒体(Sigma、MO、USA)で再構成されました。 ENPのサイズは、ホモダインダイナミクスレーザー散乱(DLS)を使用して個別に確認されました。簡単に説明すると、海水サンプルを0.22μmミリポアメンブレン(0.1N HClで事前に洗浄)で再ろ過し、5つの10 ml散乱セルに直接注ぎ、ブルックヘブンBI-200SMレーザー分光計(ブルックヘブンインスツルメンツ、ニューヨーク、米国)。 45°の角度で検出された散乱強度変動の自己相関関数は、ブルックヘブンBI 9000AT自己相関器によってオンラインで処理され、粒子サイズ分布はCONTIN法(Provencher、1982)によって計算されました。各サンプルの結果は、超音波処理の直後に3回収集されました。 DLS分光計のキャリブレーションは、単分散ラテックスミクロスフェアの標準懸濁液(Polysciences、PA、USA)を使用して実施しました。
ENP処理
植物プランクトン細胞は、L1培地を含む96ウェルプレートで24時間培養しました。細胞をENPストックで処理しました:5 mg / mlおよび1mg / ml、0.5 mg / ml、0.1 mg / ml、10μg/ ml、1μg/ mlのTiO 2 、SiO 2 、およびCeO 2 (Sigma-Aldrich、MO、USA)またはL1培地(コントロール)で48時間。分泌されたEPSを含む上清を収集し、4000 rpmで短時間遠心分離して、残留ENPを除去しました。このプロトコルは、以前の出版物[34]から採用されました。ここで使用される濃度範囲は、環境内の現在のENPレベルを表すまたは模倣することを意図したものではなく、海洋植物プランクトンに対するENPの潜在的な影響を完全に評価し、関連する細胞メカニズムを調査することを目的としています。有望な創発的ナノ材料として、ENPはまだ完全な商業的能力に達していない。 ENPがより多くのENPを海洋に導入するために商業用および家庭用製品市場に参入する前に、それらの完全な生態学的影響の詳細な評価が大いに必要です。
酵素結合レクチンアッセイ(ELLA)
分泌された多糖を含む上清を収集し、1700 rcf(Megafuge 1.0R)で短時間遠心分離して、残留ENPを除去しました。次に、上清を96ウェル(Nunc MaxiSorp、VWR、CA、USA)プレートで4℃で一晩インキュベートしました。その後、96ウェルプレートをPBST(PBS + 0.05%Tween-20)とPBSで洗浄し、1%BSAでブロックしました。 96ウェルプレートをPBSTおよびPBSで再度洗浄し、西洋ワサビペルオキシダーゼ(HRP; 5 mg / ml)(Sigma-Aldrich、MO)に結合したレクチン(Concanavalin A、ConA)(Sigma-Aldrich、MO、USA)とインキュベートしました。 、米国)、37°Cで1時間。基質である3,39,5,59-テトラメチルベンジジン(TMB; Sigma-Aldrich、MO、USA)を各ウェルに室温で添加し、続いてH 2 反応を停止するためのSO4(Sigma-Aldrich、MO、USA)。光学密度は、PerkinElmer VICTOR3(MA、USA)によって450nmで測定されました。このプロトコルは、以前の出版物[34、52]から採用されました。
DNAの決定
植物プランクトンを含むペレットを収集し、ZR-96 Quick-gDNAキット(ZYMO Research、CA、USA)を入手しました。簡単に説明すると、4×溶解バッファーを使用して植物プランクトン細胞を破壊し、DNA結合カラムを通過させ、最後に溶出バッファーで溶出しました。 DNA濃度は、NanoDrop ND-1000(Thermo、CA、USA)によって測定されました。プロトコルは、製造されたキットプロトコルから採用されました。
細胞内Ca 2+ の測定 ENPによって誘発される濃度
次に、植物プランクトン細胞にFluo-4AM色素(1 mM)(Kd =335 nM、λEx=494 nm、およびλEm=506 nm、ThermoFisher、CA、USA)を60分間ロードしました[31]。色素をロードした後、植物プランクトン細胞をすすぎ、L1培地でインキュベートし、1 mg / ml TiO 2 で処理しました。 、SiO 2 、およびCeO 2 それぞれ。すべてのカルシウムシグナル伝達実験は、Nikon顕微鏡(Nikon Eclipse TE2000-U、東京、日本)で実施されました。プロトコルと条件は、以前の出版物[31、34]から採用されました。
ENP測定のゼータ電位
ENPの表面電荷を測定するために、ENPのゼータ電位(ζ)を、25°Cの人工海水の存在下でZetasizer Nano ZS、Malvernを使用して測定しました。各サンプルからデータを収集した後、記録された値を平均しました。
統計分析
データは平均値±SDとして報告されます。各実験は、少なくとも3回独立して実行されました。ヒストグラムはGraphPadPrism6.0によって作成されました。 (GraphPad Software、Inc。、米国カリフォルニア州サンディエゴ)。
ナノマテリアル
- 半導体ナノ粒子
- プラズモンナノ粒子
- 庭からの黄色染料に対するpHの影響
- テンプレートのない表面に気相から堆積した秩序だった高密度に充填されたナノ粒子超格子の形成メカニズム
- 従来の抗生物質の殺菌効果を活性化するための排出ポンプおよびバイオフィルム阻害剤としてのナノ粒子
- 子宮頸がんにおける抗がん効果を高めるための葉酸受容体を標的としたバイオフラボノイドゲニステインをロードしたキトサンナノ粒子
- ナノテクノロジー:invivoイメージングシステムから制御されたドラッグデリバリーまで
- La1 − xSr xMnO3マンガン酸塩ナノ粒子の合成方法がそれらの特性に及ぼす影響
- クルクミンの処理効果を持つPEGコーティングされたCoFe2O4ナノ粒子の毒性
- アナターゼ-ルチル比とTiO2ナノ粒子の光触媒性能に及ぼす酸性解膠剤の影響
- 酸化還元刺激によって誘発される放出のためのナノ粒子を標的とする新規の二重ミトコンドリアおよびCD44受容体