


microRNA-342-5pのダウンレギュレーションまたはWnt3aのアップレギュレーションは、血管新生を阻害し、アテローム性動脈硬化症マウスのアテローム性動脈硬化症のプラーク安定性を維持します
要約
microRNA-342-5p(miR-342-5p)がアテローム性動脈硬化症(AS)に関与していることを示す証拠がありますが、その固有の調節メカニズムについてはほとんど知られていません。ここでは、不安定プラークの形成とASの血管新生に対するWnt3aを標的とするmiR-342-5pの効果を調査することを目的としました。 ApoE -/- AS不安定プラークモデルを複製するために、マウスに高脂肪飼料を16週間与えた。 ASの大動脈組織におけるmiR-342-5pおよびWnt3aの発現が検出されました。 miR-342-5pとWnt3aのターゲット関係を確認しました。さらに、ApoE -/- マウスにmiR-342-5pアンタゴミールと過剰発現-Wnt3aベクターを注射して、血清脂質レベル、炎症性および酸化ストレス関連サイトカイン、大動脈プラークの安定性、ASマウスのプラークにおける血管新生における機能をテストしました。 ASマウスの大動脈組織ではmiR-342-5pの発現が増強され、Wnt3aの発現が低下し、miR-342-5pはWnt3aを直接標的としました。 Wnt3aをアップレギュレートするかmiR-342-5pをダウンレギュレートすると、血中脂質含有量、炎症性および酸化ストレスレベル、大動脈組織プラークの脆弱性が減少し、ASマウスの大動脈プラークにおける血管新生が抑制されました。機能研究によると、miR-342-5pを枯渇させると、大動脈組織のプラークが安定し、Wnt3aを回復することでASマウスのプラークの血管新生が減少することが示されています。
はじめに
アテローム性動脈硬化症(AS)は、動脈の肥厚、狭窄、硬化、およびアテローム性動脈硬化症のプラークの形成を特徴とする加齢性動脈疾患です[1]。これは、先進国における一般的な死因および罹患率です[2]。ヒトのアテローム性動脈硬化症の組織病理学的研究は、プラークの発達と破裂が脂質/壊死性コアの拡大、平滑筋細胞数の減少、マクロファージ浸潤、およびコラーゲン含有量の減少によって特徴づけられることを示しています[3]。 ASの重要な細胞要素は、高脂血症、泡沫細胞の形成、マクロファージへの分化、単球の動員、および誘発された炎症で構成されています[4]。 ASの治療のための多くの薬がクリニックで広く使用されていますが、一部のサブグループの患者は依然として心筋梗塞、心筋虚血、心不全、脳卒中のリスクが高いです[5]。したがって、潜在的な分子メカニズムのさらなる調査は、AS治療のより多くの証拠を提供することができます。
単一のマイクロRNA(miRNA)は、複数の遺伝子ターゲットを同時に制御できます[6]。 miR-342-5pは、インプリントされた14q32 miRNAクラスターに含まれることが調査されており、革新的なNotch下流分子として機能し[7]、トランスフォーミング増殖因子βシグナル伝達や血管内皮増殖因子などの複数の血管新生経路を調節します[8]。 miR-342-5pなどの関連する免疫調節マイクロRNAは、アテローム性動脈硬化症の進行を調節する上で複数の重要な役割を果たします[9]。さらに、miR-155やmiR-217などの一部のmiRNAは、ASの解決に関与することが示唆されています[10、11]。ある研究では、miR-342-5pがASにおけるマクロファージ活性化の新しいモジュレーターとして機能することが報告されています[12]。別の研究では、マクロファージ由来のmiR-342-5pがASを促進し、マクロファージの炎症性刺激を増加させることが明らかになっています[13]。 Qu et al 。 鎖肛ではWnt3aの発現がmiR-342-5pによって負に調節されることを発見し[14]、miR-342-5pとWnt3aの間に標的関係があることを示しています。 Wntシグナル伝達は、胚発生時に細胞極性、細胞増殖、軸形成、アポトーシスの調節に不可欠な役割を果たします[15]。中胚葉遺伝子の重要な構成要素であるWnt3aは、胚発生において重要な役割を果たします[16]。 AS回帰中のプラークマクロファージトランスクリプトームのエピゲノムガイド分析により、Wntシグナル伝達経路の活性化が明らかになることが示されています[17]。さらに、ある研究では、Wnt3aが血管平滑筋細胞の接着と遊走を調節し、ASと再狭窄の病因に寄与することが報告されています[18]。したがって、この研究では初めて、不安定プラークの形成とASの血管新生に対するmiR-342-5pを標的としたWnt3aの効果を調査しました。
材料と方法
倫理声明
動物は、国立衛生研究所の実験動物の管理と使用に関するガイドの推奨事項に準拠した承認された手順を使用して人道的に扱われました。このプロトコルは、青海省人民病院の施設内動物管理使用委員会によって承認されました(倫理番号:201870726)。
実験動物
男性のApoE -/- 8週齢のマウスとC57BL / 6Jマウス(特定病原体除去グレード)は、Beijing Vital Laboratory Animal Technology(北京、中国)から入手できました。マウス(ケージに5〜6匹のマウス)を12時間/ 12時間の昼/夜のサイクルで飼育し、餌と水を自由に摂取させました。
ASのマウスモデルの確立
ApoE -/- AS不安定プラークモデルを確立するために、マウスに高脂肪飼料を16週間与えた。 C57BL / 6 Jマウスは、自然の飲み物と食べ物を含む通常のグループとして使用されました。 ApoE -/- マウスは12週間後に光沢のある明るい髪と背中の毛の脱落がありました。 3匹のモデルマウスの大動脈弓と腕頭動脈を解剖してヘマトキシリン-エオシン(HE)染色を行い、内膜に有意なプラーク沈着はありませんでした。別の3匹のモデル化されたマウスが4週間後に再び同定され、HE染色は大動脈弓の内膜に明らかなプラーク沈着があったことを示し、モデル確立の成功を示しています。
マウスのグループ化と治療
ApoE -/- AS不安定プラークのマウスを6つのグループに分け、各グループに12匹のマウスを入れました:ASグループ、ネガティブコントロール(NC)グループ(ApoE -/- に生理食塩水を注射) マウス)、miR-342-5pアゴミールグループ(ApoEでmiR-342-5p発現を過剰発現させるためにmiR-342-5pアゴミールを注射 -/- マウス)、miR-342-5pアンタゴミールグループ(ApoE -/- でのmiR-342-5p発現を低下させるためにmiR-342-5pアンタゴミールを注射) マウス)、過剰発現(oe)-Wnt3aグループ(ApoE -/- でWnt3a発現をアップレギュレートするためにoe-Wnt3aベクターを注入 マウス)およびmiR-342-5pアゴミール+ oe-Wnt3aグループ(miR-342-5pアゴミールおよびoe-Wnt3aベクターを注射して、ApoE -/-<におけるmiR-342-5pおよびWnt3aの発現をアップレギュレートします。 / sup> マウス)。通常のグループとしてのC57BL / 6Jマウスには通常の食餌を与えました。高脂肪飼料には、20%の脂肪と0.25%のコレステロールが含まれていました。 miR-342-5pアゴミール、miR-342-5pアンタゴミール、oe-Wnt3aベクターは、Sangon(上海、中国)から購入しました。 oe-Wnt3aベクター、miR-342-5pアゴミール、miR-342-5pアンタゴミールはすべて、0.2 mLの生理食塩水に溶解し、2週間ごとに尾静脈から40 mg / kgの用量でマウスに注射しました。 8週間後、眼球から血液サンプルを採取し、マウスを安楽死させて動脈組織を収集しました[19]。
予備実験では、ApoE -/- ASのマウスには、10 mg / kg、20 mg / kg、40 mg / kgのmiR-342-5pアゴミール、miR-342-5pアンタゴミール、またはoe-Wnt3aベクターを注射しました(2週間に1回、合計4回)。次に、β-カテニンの発現レベルを逆転写定量的ポリメラーゼ連鎖反応(RT-qPCR)によって検出しました。
サンプルの収集と処理
サンプリングする前に、マウスを12時間絶食させ、エーテル吸入で麻酔をかけ、眼球から血液サンプルを採取しました。マウスの胸部を開き、胸部大動脈を腹部大動脈の端まで解離させ、血管全体を取り出した。 RNAを含まないリン酸緩衝生理食塩水(PBS)で洗浄した後、組織を包埋してHE染色、オイルレッドO、シリウスレッド染色、免疫組織化学的染色を行いました。一部の維管束組織は、RT-qPCRおよびウエスタンブロットのために-80°Cで保存されました。
血中脂質レベルの検出
自動生化学分析装置(Roche、バーゼル、スイス)を採用して、血清中の総コレステロール(TC)、トリグリセリド(TG)、低密度リポタンパク質コレステロール(LDL-C)、および高密度リポタンパク質コレステロール(HDL-C)を検出しました。検出は、キットの仕様に従って実装されました(NanJing JianCheng Bioengineering Institute、Nanjing、China)。
酵素結合免疫吸着測定法(ELISA)
血清サイトカイン含有量の測定:市販のインターロイキン(IL)-5、IL-12p70、腫瘍壊死因子α(TNF-α)およびインターフェロン(IFN)-γELISAキットを使用しました。最後に、各ウェルの光学密度(OD)値を、マイクロプレートリーダーで450nmでテストしました。
酸化ストレス損傷の決定:血清中のマロンジアルデヒド(MDA)含有量とスーパーオキシドジスムターゼ(SOD)活性は、MDAキット(OD値は532 nmで分光光度計によってテストされました)およびSODキット(OD値は450でマイクロプレートリーダーによって決定されました)によってテストされましたnm)。 IL-5、IL-12p70、TNF-α、IFN-γ、MDA、およびSOD ELISAキットは、MultiSciences(Lianke)Biotechnology Corporate Limited(杭州、浙江省、中国)から購入しました。
HE染色、オイルレッドO染色、シリウスレッド染色
固定と埋め込みの後、標本は4マイクロメートルの厚さの連続したセクションにスライスされました。スライスを脱ロウおよび水和し、ヘマトキシリンおよびエオシンで染色し、分化させ、脱水し、キシレンで透明にし、乾燥させ、中性ガムで密封した。核は青色で、細胞質や結合組織などの他の組織はさまざまな色合いで赤色でした。プラークの形成を蛍光顕微鏡で観察した。 HE染色切片の動脈壁を顕微鏡下で選択し、実験結果をデジタルカメラで収集した。 Image Pro Plus6.0(IPP6.0)画像解析ソフトウェアモジュールを使用して、各スライスの断面のプラーク面積と壁の面積、およびそれらの比率を計算しました。
オイルレッドO染色には、4〜5μmのスライスを選択しました。スライスを過剰温度で20分間乾燥し、100%イソプロパノールで5分間インキュベートしました。次に、スライスを60°Cのオーブンで0.5%オイルレッドO染色液と8分間インキュベートし、85%イソプロパノールで3分間洗浄し、ヘマトキシリンで1分間染色し、洗浄して密封しました。オイルレッドO染色の結果は、脂質が赤またはオレンジであり、核が水色であることを示唆していました。 IPP6.0ソフトウェアを使用して、組織スライスのプラークの脂肪領域とプラーク領域を計算しました。脂質含有量=オイルレッドO陽性染色領域/プラーク領域×100%。
シリウスレッド染色:スライスを脱ロウおよび水和し、セレスティンブルー染色液で10分間染色し、シリウスレッド染色液で20分間染色し、ヘマトキシリンで10分間対比染色しました。最後に、スライスを勾配エタノールで脱水し、キシレンで透明にし、中性ガムで密封しました。組織スライスのプラークのコラーゲン面積は、IPP6.0ソフトウェアによって計算されました。コラーゲン領域=シリウスレッド陽性染色領域/プラーク領域×100%。プラーク領域の脂質とコラーゲンの割合を計算しました。
ヘマトキシリン、エオシン、およびシリウス染料は、China Pharmaceutical Group Shanghai Chemical Reagent Co. Ltd.(Shanghai、China)から入手できました。オイルレッドO粉末は、Sigma-Aldrich Chemical Company(セントルイス、ミズーリ州、米国)から購入しました。
逆転写定量的ポリメラーゼ連鎖反応(RT-qPCR)
大動脈組織をトータルRNA抽出試薬Trizol(Invitrogen、Carlsbad、California、USA)に加え、ホモジナイズしてトータルRNAを抽出し、相補DNAを構成しました。プライマーはすべてGenbankが提供する配列を参照し、Primer 5.0が設計し、Shanghai Sangon Biotechnology Co. Ltdが合成しました。MiR-342-5p:フォワード:5'-CGGAGGGGTGCTATCTGTGATTGAG-3 '、リバースプライマーはキットユニバーサルプライマー(Qiagen会社、ヒルデン、ドイツ); Wnt3a:順方向:5'-AGGTAAGCTACTCCCTCAACTA-3 '、逆方向:5'-CTGAAGCACCCTCTCATGTATC-3'; β-アクチン:順方向:5'-GCACCACACCTTCTACAATGAGC -3 '、逆方向:5'-TCGTTGCCAATAGTGATGACC-3'; β-カテニン:順方向:5'-TCAAGAGAGCAAGCTCATCATTCT-3 '、逆方向:5'-CACCTTCAGCACTCTGCTTGTG-3'。反応後、閾値サイクル(Ct)をコンピューターで分析した。 miR-342-5pとU6の相対比をその発現として使用し、Wnt3aとβ-アクチンの相対比をその発現として使用し、相対比を2 -ΔΔCt で計算しました。 メソッド。
ウエスタンブロット分析
総タンパク質は大動脈組織から抽出されました。タンパク質濃度はビシンコニン酸法で測定した。ポリアクリルアミドゲル電気泳動を行った。次に、タンパク質をポリフッ化ビニリデンメンブレンに転写し、ターゲットバンドを取得しました。メンブレンを5%スキムミルクで1時間密封し、一次抗体Wnt3a(1:500)、β-カテニン(1:1000、Santa Cruz Biotechnology、Inc.、Santa Cruz、CA、USA)、CD34(1 :2500、米国カリフォルニア州アブカム)およびβ-アクチン(1:2000、Beyotime Biotechnology Co.、上海、中国)、4°Cで一晩。メンブレンをTween20(pH =7.5、10 mmol / L Tris–HCl、100 mmol / L NaCl、0.2%Tween-20)を含むトリス緩衝生理食塩水で10分×3回洗浄した後、二次抗体を添加しました。 (1:1000、ZSGB-Bio、北京、中国)2時間。バンドのグレー値を評価し、タンパク質発現を定量化するために、ImageJソフトウェアが採用されました。
免疫組織化学的染色
4〜5 µmのスライスを、100 mg / Lのポリリジンでコーティングし、アセトンで固定したスライドに配置しました。内因性ペルオキシダーゼはウシ血清アルブミンによってブロックされました。組織にMOMA-2抗体(1:200)、α-SMA(1:200)、CD34(1:200、Abcam Inc.、米国マサチューセッツ州ケンブリッジ)を滴下し、二次抗体作用溶液(1:200)を添加しました。 1:1000)。組織はジアミノベンジジンによって発達し、ヘマトキシリンによって対比染色され(1分)、脱水され、透明にされ、密封され、顕微鏡で観察されました。免疫組織化学的セクションごとに3つの異なる視野が選択されました。定量分析のためにIPP6.0ソフトウェアを実行しました。それぞれMOMA-2およびα-SMAの陽性免疫組織化学的染色は、マクロファージおよび平滑筋細胞が主に黄色から茶色の細胞質に位置していることを示しています。マクロファージと平滑筋細胞のパーセンテージを別々に計算し、プラーク中の脂質とコラーゲンのパーセンテージと組み合わせて、プラーク脆弱性指数を計算しました。プラークの脆弱性指数=(マクロファージの陽性率+脂質の陽性率)/(コラーゲンの陽性率+平滑筋細胞の陽性率)[20]。微小血管密度(MVD)は、CD34発現の測定によって評価され、微小血管の数/ mm 2 として定量化されました。 。
デュアルルシフェラーゼレポーター遺伝子アッセイ
miR-342-5pの標的遺伝子は、生物学的予測Webサイト(http://www.microRNA.org)によって分析されました。デュアルルシフェラーゼレポーター遺伝子アッセイを使用して、Wnt3aがmiR-342-5pの標的遺伝子であるかどうかを確認しました。 Wnt3a 3 '非翻訳領域(3'-UTR)の野生型または変異型配列を、GP-miRGLOベクター(GenePharma、上海、中国)にクローニングしました。レポーター(0.5μg)と1、10、または100 pM miR-342-5pアゴミールをマウス大動脈内皮細胞(No. 506、MingzhouBio、寧波、中国)に48時間トランスフェクトし、デュアルルシフェラーゼアッセイシステムを使用してルシフェラーゼ活性をテストしました。 (Promega、WI、USA)。
統計分析
すべてのデータは、SPSS 21.0ソフトウェア(IBM Corp. Armonk、NY、USA)によって解釈されました。測定データは平均±標準偏差として示されました。 2つのグループ間の格差は t によって定式化されました -一元配置分散分析(ANOVA)とそれに続くテューキーの多重比較検定による複数のグループ間の検定。統計的有意性は P によって確立されました 値<0.05。
結果
miR-342-5pはApoEの大動脈組織で増加およびWnt3aは減少します -/- マウスとmiR-342-5pはWnt3aを直接標的にします
マイクロRNA(miRNA)の標的遺伝子は、アテローム性動脈硬化症に関連する機能に関連しています。 miR-342-5p、Wnt3aおよびβ-カテニンは、RT-qPCRおよびウエスタンブロットアッセイによってASモデルマウスの大動脈組織でテストされました。正常群と比較して、AS群ではmiR-342-5pが上昇し、Wnt3aとβ-カテニンが低下していることが明らかになりました( P <0.05)。 NCグループと比較して、miR-342-5pは増強され、Wnt3aとβ-カテニンはmiR-342-5pアゴミールグループ(両方 P )で減少しました。 <0.05)、miR-342-5pは減少しましたが、Wnt3aとβ-カテニンはmiR-342-5pアンタゴミールグループ(両方 P )で上昇しました。 <0.05)。 Wnt3aとβ-カテニンの発現は、NCグループと比較してoe-Wnt3aグループで高くなりました(両方の P <0.05)。 miR-342-5pアゴミールグループと比較して、Wnt3aおよびβ-カテニンの発現は、miR-342-5pアゴミール+ oe-Wnt3aグループで上昇しました( P <0.05)(図1A–D)。さらに、予備実験では、異なる濃度のmiR-342-5pアゴミール、miR-342-5pアンタゴミール、およびoe-Wnt3aの処理下でのβ-カテニン発現がテストされ、結果が示されました(追加ファイル1:図S1 )miR-342-5pアゴミール濃度が高いほど、β-カテニンの発現は低くなります。 miR-342-5pアンタゴミール濃度が高いほど、β-カテニンの発現が高くなります。 oe-Wnt3a濃度が高いほど、β-カテニンの発現が高くなります。
ApoE -/- の大動脈組織では、miR-342-5pが増加し、Wnt3aが減少します。 マウスとmiR-342-5pはWnt3aを直接標的とします。 A RT-qPCRによって検出されたマウスの大動脈組織におけるmiR-342-5pの発現。 B RT-qPCRによって検出されたマウスの大動脈組織におけるWnt3amRNAの発現( n =12)。 C 、 D ウエスタンブロット分析によってテストされたマウスの大動脈組織におけるWnt3aおよびβ-カテニンタンパク質の発現( n =12)。 E Wnt3a3'-UTR内のmiR-342-5pの結合部位。 F miR-342-5pアゴミールは、Wnt3a 3'-UTR( N )でトランスフェクトされた細胞の相対活性を用量依存的に減少させました。 =3)。 G miR-342-5pアゴミールまたはスクランブル( N )でトランスフェクトした後の野生型および変異型Wnt3a3'-UTRを含む細胞におけるルシフェラーゼの相対的活性 =3)。 * P <0.05対通常のグループ、 # P <0.05対NCグループ。 & P <0.05対miR-342-5pアゴミールグループ。測定データは平均±標準偏差として示されました。 2つのグループ間の比較は、 t によって作成されました。 -テスト。複数のグループ間の比較は、一元配置分散分析とそれに続くテューキーの多重比較検定によって評価されました。 AS、アテローム性動脈硬化症; NC、ネガティブコントロール
miRNAは、メッセンジャーRNA 3'UTRに結合することにより、特定の遺伝子の翻訳を阻害する可能性があります。バイオインフォマティクスのウェブサイトは、miR-342-5pとWnt3aの間にターゲット関係があると予測しました(図1E)。デュアルルシフェラーゼレポーター遺伝子アッセイは、Wnt3a 3'UTRベクターでトランスフェクトされたマウス大動脈内皮細胞において、ルシフェラーゼのレニラ/ホタル値がmiR-342-5pアゴミールのトランスフェクションによって用量依存的に減少し、10から100に有意に減少したことを報告しましたNCグループと比較した場合、100 pM miR-342-5pアゴミールグループでは、pM miR-342-5pアゴミールと64%の減少が発生しました。これは、Wnt3a3'UTRにmiR-342-5p標的部位が存在することを示しています。ただし、ルシフェラーゼ活性のレニラ/ホタル値は、Wnt3a変異グループでは影響を受けませんでした(図1F、G)。したがって、Wnt3aはmiR-342-5pの直接の標的遺伝子であり、miR-342-5p / Wnt3aはASの進行を調節できることが確認できました。
ApoEの脂質レベルに対するアップレギュレーションされたWnt3aまたはダウンレギュレーションされたmiR-342-5pの影響 -/- マウス
さらに、miR-342-5pがWnt3aシグナル伝達経路を標的にして調節することが、ASマウスの脂質レベルに影響を与えるかどうかを調べるために、自動生化学的分析装置を利用して脂質レベルの変化を観察しました。その結果、正常群とは対照的に(図2A〜D)、AS群(すべて P )でTC、TG、LDL-Cの含有量が増加し、HDL-Cの含有量が減少したことが明らかになりました。 <0.05)。 NCグループと比較して、miR-342-5pアゴミールグループ(すべて P )では、TC、TG、およびLDL-Cの含有量が増加し、HDL-Cの含有量が減少しました。 <0.05)、TC、TG、LDL-Cの含有量は減少し、HDL-Cの含有量はmiR-342-5pアンタゴミールグループとoe-Wnt3aグループ(すべて P )で増加しました。 <0.05)。 miR-342-5pアゴミールグループに関連して、TC、TG、LDL-Cの含有量が減少し、HDL-Cの含有量がmiR-342-5pアゴミール+ oe-Wnt3aグループ(すべて P <0.05)。これらの結果は、miR-342-5pとWnt3aがASマウスの血中脂質レベルを調節することを示唆し、miR-342-5pとWnt3aの間の標的調節関係をさらに示しました。 Wnt3aの過剰発現は、ASマウスに対する過剰発現したmiR-342-5p誘発効果を逆転させます。
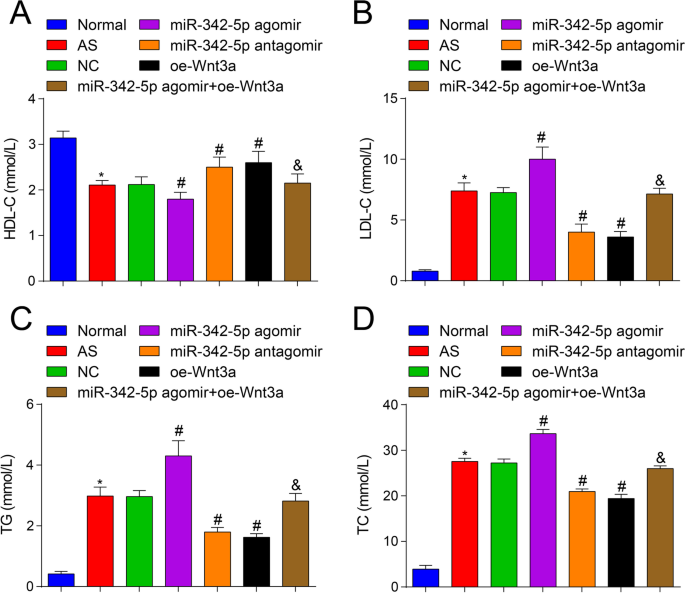
ApoEの脂質レベルに対するアップレギュレーションされたWnt3aまたはダウンレギュレーションされたmiR-342-5pの効果 -/- マウス。 A マウス群の血清中のHDL-C含有量の比較。 B マウス群の血清中のLDL-C含有量の比較。 C マウス群の血清中のTG含有量の比較。 D マウス群の血清中のTC含有量の比較。 n =12. * P <0.05対通常のグループ、 # P <0.05対NCグループ。 & P <0.05対miR-342-5pアゴミールグループ。測定データは平均±標準偏差として示されました。複数のグループ間の比較は、一元配置分散分析とそれに続くテューキーの多重比較検定によって評価されました。 AS、アテローム性動脈硬化症; NC、ネガティブコントロール
ApoEの血清中の炎症性および酸化ストレス関連サイトカインに対するWnt3aの過剰発現またはmiR-342-5pの低発現の影響 -/- マウス
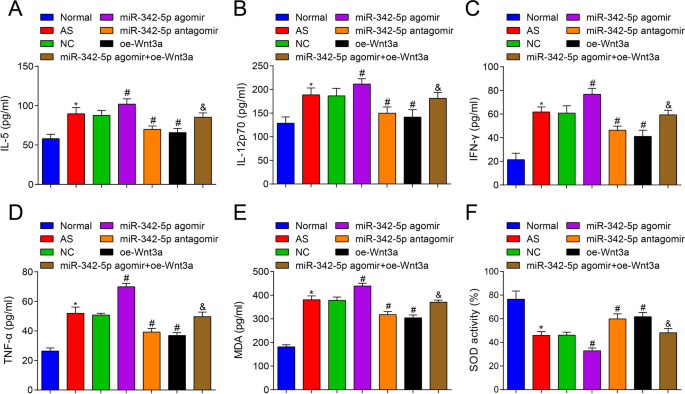
次に、ASマウスの血清中のサイトカインの含有量をELISAでテストした結果、正常群(IL-5、IL-12p70、IFN-γ、TNF-α)と比較して(図3A–D)が増強されたことが報告されました。 ASグループ内(すべての P <0.05)。 NCグループと比較して、IL-5、IL-12p70、IFN-γ、およびTNF-αの含有量は、miR-342-5pアゴミールグループ(すべて P )で上昇しました。 <0.05)、IL-5、IL-12p70、IFN-γおよびTNF-αの含有量は、miR-342-5pアンタゴミールグループおよびoe-Wnt3aグループ(すべて P )で分解されました。 <0.05)。 miR-342-5pアゴミールグループとは対照的に、IL-5、IL-12p70、IFN-γおよびTNF-αの含有量は、miR-342-5pアゴミール+ oe-Wnt3aグループで減少しました(すべて P <0.05)。 miR-342-5pが標的とするWnt3aシグナル伝達経路の調節は、ASマウスの血清中の関連サイトカインのレベルをさらに調節することが示唆されました。
ApoEの血清中の炎症性および酸化ストレス関連サイトカインに対するWnt3aの過剰発現またはmiR-342-5pの低発現の影響 -/- マウス。 A マウス群の血清中のIL-5含有量の比較。 B マウス群の血清中のIL-12p70含有量の比較。 C マウス群の血清中のIFN-γ含有量の比較。 D マウス群の血清中のTNF-α含有量の比較。 E、マウス群の血清中のMDA含有量の比較。 F マウス群の血清中のSOD活性の比較。 n =12. * P <0.05対通常のグループ、 # P <0.05対NCグループ。 & P <0.05対miR-342-5pアゴミールグループ。測定データは平均±標準偏差として示されました。複数のグループ間の比較は、一元配置分散分析とそれに続くテューキーの多重比較検定によって評価されました。 AS、アテローム性動脈硬化症; NC、ネガティブコントロール
さらに、マウスの血清中のMDA含有量とSOD活性を試験したところ、正常群と比較して(図3E、F)、AS群ではMDA含有量が上昇し、SOD活性が低下していることが明らかになりました(両方 P <0.05)。 NCグループと比較して、miR-342-5pアゴミールグループ( P の両方)では、MDA含有量が増加し、SOD活性が減少しました。 <0.05)、一方、miR-342-5pアンタゴミールグループとoe-Wnt3aグループ(すべて P )では、MDA含有量が減少し、SOD活性が増加しました。 <0.05)。 miR-342-5pアゴミールグループと比較して、miR-342-5pアゴミール+ oe-Wnt3aグループ(両方 P )では、MDA含有量が低下し、SOD活性が増強されました。 <0.05)。したがって、miR-342-5pを枯渇させ、Wnt3aを回復させると、ASマウスの酸化ストレスが抑制されるという要約が得られました。
miR-342-5pの枯渇またはWnt3aの回復がApoEの大動脈プラークの脂質およびコラーゲン含有量に及ぼす影響 -/- マウス
miR-342-5pを標的としたWnt3aがマウスの大動脈組織のプラーク領域に及ぼす影響を調べるために、HE染色を行った結果、正常群を除いて(図4A、B)、すべてのASプラークが形成されたことが明らかになりました。他のグループのセクション。 AS群では、プラーク面積が大きく、線維性被膜が薄く、脂質コアが拡大し、泡沫細胞とコレステロール結晶の沈殿がプラークに多く現れ、動脈の内壁と筋肉層が厚くなり、プラークは不安定。 NCグループの状況はASグループの状況と同様でした。 miR-342-5p antagomirおよびoe-Wnt3aグループでは、プラーク面積が小さく、動脈の内膜が滑らかで、線維性被膜の数が少なく、薄くなりました。破裂はありませんでしたが、プラークにはさまざまなサイズの泡沫細胞がありました。コレステロール結晶は非対称に分布し、部分的に石灰化しており、平滑筋細胞とコラーゲン線維の数が増加し、プラークは安定する傾向がありました。 NC群と比較して、miR-342-5pアゴミール群ではプラーク面積が増加し、AS病変が悪化した( P <0.05)一方、プラーク面積はmiR-342-5pアンタゴミールグループとoe-Wnt3aグループで減少し、AS病変が減少しました(両方 P <0.05)。 miR-342-5pアゴミールグループと比較して、プラーク面積はmiR-342-5pアゴミール+ oe-Wnt3aグループで減少しました( P <0.05)。
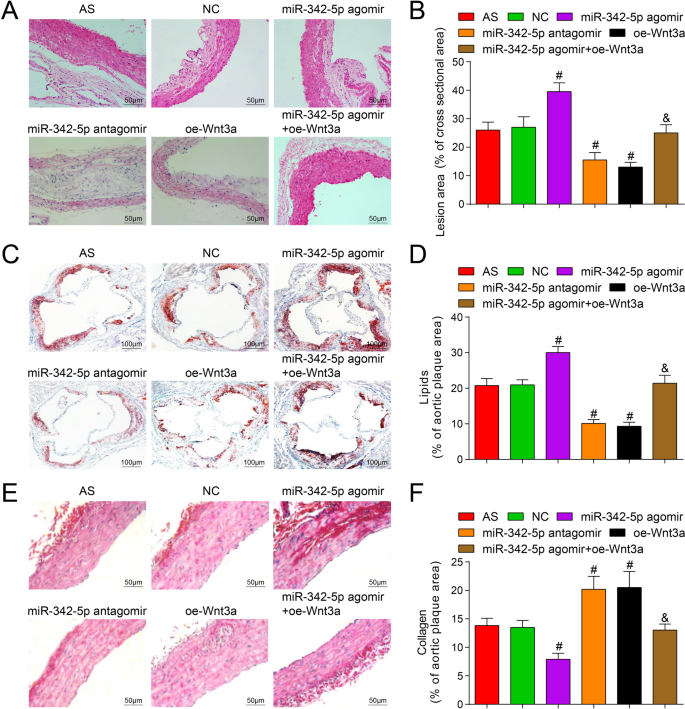
miR-342-5pの枯渇またはWnt3aの回復が、ApoEの大動脈プラークの脂質およびコラーゲン含有量に及ぼす影響 -/- マウス。 A マウスの大動脈HE染色の結果(スケールバー:50μm)。 B マウスの各グループにおける大動脈プラーク面積の比較。 C マウスのすべてのグループでの大動脈オイルレッドO染色の結果(スケールバー:100μm)。 D マウスの大動脈組織における脂質含有量の比較。 E マウスの各グループにおける大動脈シリウスレッド染色の結果(スケールバー:50μm)。 F マウスの大動脈組織におけるコラーゲン含有量の比較。 n =12. # P <0.05対NCグループ。 & P <0.05対miR-342-5pアゴミールグループ。測定データは平均±標準偏差として示されました。複数のグループ間の比較は、一元配置分散分析とそれに続くテューキーの多重比較検定によって評価されました。 AS、アテローム性動脈硬化症; NC、ネガティブコントロール
オイルレッドO染色とシリウスレッド染色を採用して、miR-342-5pを標的としたWnt3aがマウスの大動脈組織のプラークの脂質含有量とコラーゲン含有量に及ぼす影響を検出し、その結果は次のことを示しました(図4C–F)オイルレッドO染色は赤い脂肪と青い核を示したが、シリウスレッド染色は赤いコラーゲン線維と青い核を示した。 NCグループと比較して、miR-342-5pアゴミールグループでは脂質含有量が増加し、コラーゲン含有量が減少し、miR-342-5pアンタゴミールグループとoe-Wnt3aでは脂質含有量が減少し、コラーゲン含有量が蓄積されました。グループ(すべて P <0.05)。 miR-342-5pアゴミールグループでは、miR-342-5pアゴミール+ oe-Wnt3aグループ(両方 P )で脂質含有量が減少し、コラーゲン含有量が増加しました。 <0.05)。実験結果は、miR-342-5pが標的とするWnt3aシグナル伝達経路の調節が、ASマウスの大動脈プラークの脂質およびコラーゲン含有量に調節効果を及ぼすことを完全に示しています。
ApoEの大動脈プラークのマクロファージおよび平滑筋細胞に対するダウンレギュレーションされたmiR-342-5pまたはアップレギュレーションされたWnt3aの効果 -/- マウス
ASの程度は単核マクロファージの含有量に正比例します[21]。 VSMCは動脈の中間層の主要な細胞であり、動脈壁の完全性を維持するために不可欠です。 VSMCは動脈壁の再建に関与し、さまざまな段階でASで重要な役割を果たします[22]。 α-SMAは平滑筋細胞の特異的マーカーです[23]。この研究では、マクロファージマーカー抗体(MOMA-2)を使用してマクロファージを標識し、免疫組織化学を適用して、それぞれMOMA-2とα-SMAの発現を検出しました。
顕微鏡下で、MOMA-2およびα-SMAのそれぞれの陽性免疫組織化学的染色は、マクロファージおよび平滑筋細胞が主に黄色から茶色の細胞質に位置していることを示しています。 MOMA-2免疫陽性は、マクロファージが主に黄色から茶色の細胞質に存在することを示しました。免疫組織化学によって決定された、NCグループと比較して、プラークマクロファージ(MAMO-2)陽性染色のパーセンテージが上昇し、陽性平滑筋細胞のパーセンテージがmiR-342-5pアテロームで減少したことが明らかになった(両方 P <0.05)。 miR-342-5pアンタゴミールグループとoe-Wnt3aグループ(すべて P )では、プラークマクロファージ(MAMO-2)陽性染色の割合が減少し、陽性平滑筋細胞の割合が増加しました。 <0.05)。 miR-342-5pアテロームグループと比較して、プラークマクロファージ(MAMO-2)陽性染色の割合は低下し、陽性平滑筋細胞の割合はmiR-342-5pアテローム+ oe-Wnt3aグループで上昇しました(両方 P <0.05)(図5A–D)。 miR-342-5pが標的とするWnt3aシグナル伝達経路の調節は、ASマウスの動脈組織プラークにおけるマクロファージと平滑筋細胞の凝集を調節できることが示唆されました。
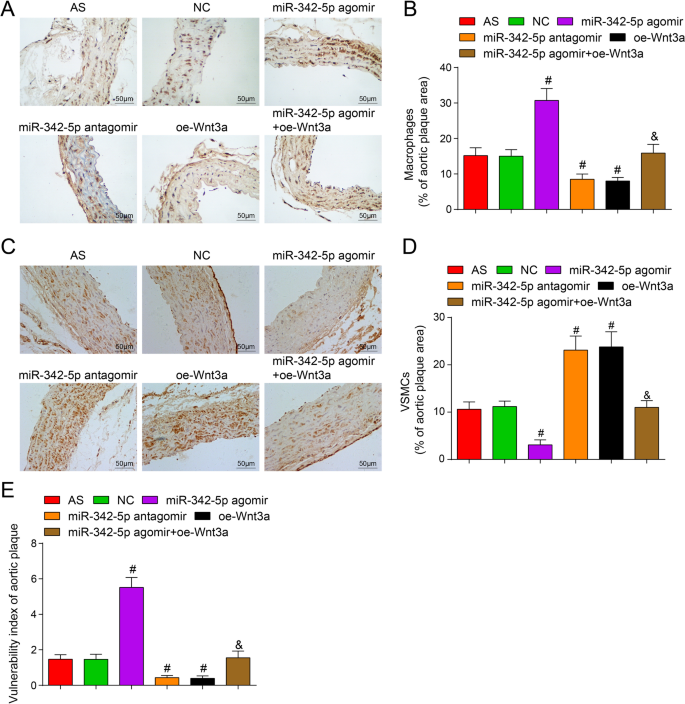
ApoEの大動脈プラークの脆弱性に対するWnt3aの高発現またはmiR-342-5pの低発現の影響 -/- マウス。 A マウスの各グループにおけるMOMA-2の免疫組織化学的染色(スケールバー:50μm)。 B 図Aの定量分析。 C マウスの各グループにおけるα-SMAの免疫組織化学的染色(スケールバー:50μm)。 D 図 C の定量分析 。 E ASマウスの大動脈組織におけるプラーク脆弱性指数の比較。 n =12. # P <0.05対NCグループ。 & P <0.05対miR-342-5pアゴミールグループ。測定データは平均±標準偏差として示されました。複数のグループ間の比較は、一元配置分散分析とそれに続くテューキーの多重比較検定によって評価されました。 AS、アテローム性動脈硬化症; NC、ネガティブコントロール
ApoEの大動脈プラーク脆弱性に対するWnt3aの高発現またはmiR-342-5pの低発現の影響 -/- マウス
プラーク脆弱性指数を計算した:(マクロファージの陽性パーセンテージ+脂質の陽性パーセンテージ)/(平滑筋細胞の陽性パーセンテージ+コラーゲンの陽性パーセンテージ)。 NCグループに関連して、プラーク脆弱性指数はmiR-342-5pアゴミールグループで上昇しました( P <0.05)、miR-342-5pアンタゴミールグループとoe-Wnt3aグループ(両方 P )で減少しました <0.05)。 miR-342-5pアゴミールグループと比較して、プラーク脆弱性指数はmiR-342-5pアゴミール+ oe-Wnt3aグループで減少しました( P <0.05)(図5E)。簡単に説明すると、miR-342-5pは、ASマウスの動脈組織におけるプラークのWnt3aシグナル伝達経路を介した脆弱性の調節を標的としました。
ApoEの大動脈プラークにおける血管新生に対するmiR-342-5pの低発現またはWnt3aの過剰発現の影響 -/- マウス
内皮細胞マーカーCD34に対する抗体は、血管密度を検出できます[24]。免疫組織化学およびウエスタンブロットによる。 NCグループと比較して、MVDはmiR-342-5pアゴミールグループで上昇し、miR-342-5pアンタゴミールグループとoe-Wnt3aグループで減衰しました(すべて P <0.05)。 miR-342-5pアゴミールグループと比較して、MVDはmiR-342-5pアゴミール+ oe-Wnt3aグループで減少しました( P <0.05)(図6A–C)。まとめると、Wnt3aシグナル伝達経路を標的とし調節するmiR-342-5pは、ASマウスのプラークにおける血管新生の密度の調節に直接関与しました。
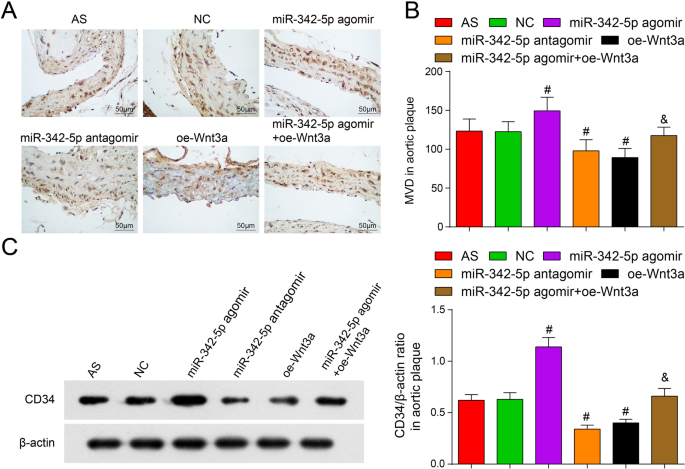
ApoEの大動脈プラークにおけるMVDに対するmiR-342-5pの低発現またはWnt3aの過剰発現の影響 -/- マウス。 A ApoEの各グループにおけるCD34の免疫組織化学的染色 -/- マウス(スケールバー:50μm)。 B ApoEの大動脈プラークにおけるMVDの比較 -/- マウス。 C ApoEにおけるCD34タンパク質発現の比較 -/- マウス。 n =12. # P <0.05対NCグループ。 & P <0.05対miR-342-5pアゴミールグループ。測定データは平均±標準偏差として示されました。複数のグループ間の比較は、一元配置分散分析とそれに続くテューキーの多重比較検定によって評価されました。 AS、アテローム性動脈硬化症; NC、ネガティブコントロール
ディスカッション
ASは、慢性炎症と血管リモデリングの形態を伴う予測不可能な疾患であり、世界的な死亡率と罹患率の主な原因です[25]。以前の研究では、miR-342-5pがASの進行の調節に関与していることが議論されています[26]。また、Wnt経路が糖尿病性ASの発生と発症の促進に関与していることも反映されています[27]。 ASにおけるmiR-342-5pとWnt3aの関連するメカニズムはまだ発掘されていないため、私たちの研究は、不安定プラークの形成とASの血管新生に対するmiR-342-5pを標的としたWnt3aの効果を調査することでした。
私たちの研究では、ASマウスの大動脈組織で高発現のmiR-342-5pと低発現のWnt3aが見つかったことが明らかになりました。ある研究によると、マクロファージ由来のmiR-342-5pは、ApoEの初期のアテローム性動脈硬化症の病変で劇的に上昇します -/- マウス[13]。別の研究では、miR-342-5pが心房細動患者で著しく上昇していることが示されています[28]。 Wnt3aの欠損は、造血幹細胞の自己複製を不可逆的に損傷し、前駆細胞の分化に欠陥をもたらすことが報告されています[29]。ある研究では、Wnt3aの枯渇が心臓機能の欠陥につながることが示されています[30]。アルツハイマー病マウスの海馬におけるWnt3aの発現が著しく低下していることが明らかになっています[31]。私たちの研究からの別の結果は、Wnt3aがASマウスのmiR-342-5pによって直接標的にされたことです。 miR-342-5pはWnt3aの3'-UTRを標的とし、その発現を負に調節できることが報告されています[14]。
さらに、我々の研究は、TC、TG、LDL-C、IL-5、IL-12p70、IFN-γ、TNF-αおよびMDA含有量が血清中で増加し、HDL-C含有量およびSOD活性が減少したことを示唆しました。さらに、プラーク面積、脂質含有量、コラーゲン含有量およびMVDが増強され、MOMA-2発現が上昇し、α-SMA発現がASマウスで減少した。 IFN-γは、抗線維症、抗増殖、免疫調節、アポトーシス、抗ウイルス活性など、多くの機能を備えた可溶性サイトカインです[32]。グルタミン治療は、アルツハイマー病のSOD活性を著しく上昇させ、MDA含有量を低下させ、Wnt3aタンパク質レベルを上昇させることが明らかになっています[31]。ある研究では、TC、TG、LDL-Cの血漿レベルが著しく上昇し、HDL-CがASで著しく低下することが明らかになっています[33]。ある研究によると、代謝障害を抑制し、アテローム性動脈硬化症のプラークの安定性を高めるのに価値のあるTEMPOLサプリメントは、プラークコラーゲン含有量を高め、脂質含有量を減らします[34]。周ら。 OPCRR治療は、TC、TG、LDL-Cを含む血清脂質プロファイルを劇的に低下させ、HDL-Cを上昇させ、脂質過酸化の産物としてMDA含有量を低下させ、さらに、TNF-αの血清レベルを低下させることに注目しました。 AS [35]。アテローム性動脈硬化症のサンプルは明らかにα-SMAの発現を低下させていることが示されています[36]。ある研究では、MVDの上昇は、罹患した大動脈、特に破裂したアテローム性動脈硬化症のプラークに見られることが示されています[37]。さらに、私たちの研究では、miR-342-5pの発現不良とWnt3aの過剰発現により、脂質レベル、サイトカイン含有量、酸化ストレス応答、プラーク面積、脂質含有量が減少し、コラーゲン含有量が増加し、MOMA-2発現が減少し、αが回復したことが明らかになりました。 -ASマウスの大動脈組織におけるSMA発現。 miR-342-5pは、LDL-CおよびTNF-αの血清レベルと正の相関があり、冠状動脈疾患(CAD)患者のHDL-Cと逆相関があることが以前に示唆されています[12]。別の研究では、miR-342-5pの枯渇がASを阻害することが確認されています[13]。さらに、実験により、Wnt1の低血清レベルは、未熟なCAD患者のTGおよびLDL-Cの上昇に関連していることが示されています[38]。さらに、ある研究では、パーキンソン病ラットのクルクミン群で、Wnt3aのアップレギュレーション、SOD含有量の増加、MDA含有量の減少が見られることが示されています[39]。
結論
簡単に言えば、私たちの研究は初めてASのmiR-342-5p / Wnt3軸のメカニズムを発見し、miR-342-5pを枯渇させると、Wnt3aを回復することにより、ASマウスの不安定プラークの形成と血管新生を減少させる可能性があることを明らかにしました。 ASの治療の潜在的な候補(追加ファイル2:図S2)。 miR-342-5pは、アテローム性動脈硬化症において他のmiRNAと相乗効果を発揮する可能性がありますが、時間と資金の制約により、これ以上の関連する議論は行いませんでした。これもこの研究の限界です。
データと資料の可用性
研究で提示された元の貢献は記事/補足資料に含まれており、さらに問い合わせは対応する著者に向けることができます。
略語
- miR-342-5p:
-
MicroRNA-342-5p
- A:
-
アテローム性動脈硬化症
- α-SMA:
-
α-平滑筋アクチン
- MVD:
-
微小血管密度
- miRNA:
-
MicroRNA
- oe:
-
過剰発現
- NC:
-
ネガティブコントロール
- PBS:
-
リン酸緩衝生理食塩水
- TC:
-
総コレステロール
- TG:
-
トリグリセリド
- LDL-C:
-
低密度リポタンパク質コレステロール
- HDL-C:
-
高密度リポタンパク質コレステロール
- ELISA:
-
酵素免疫測定法
- IL:
-
インターロイキン
- TNF-α:
-
腫瘍壊死因子アルファ
- IFN:
-
インターフェロン
- MDA:
-
Malondialdeyde
- SOD:
-
スーパーオキシドジスムターゼ
- RT-qPCR:
-
逆転写定量的ポリメラーゼ連鎖反応
ナノマテリアル
- 水晶振動子の周波数偏差の特性評価:周波数耐性、周波数安定性、および経年劣化
- 状態監視が向上し、製紙の効率が維持されます
- シクロカーボンの作成とイメージング
- スピーカーとイヤホンのグラフェン
- microRNA-342-5pのダウンレギュレーションまたはWnt3aのアップレギュレーションは、血管新生を阻害し、アテローム性動脈硬化症マウスのアテローム性動脈硬化症のプラーク安定性を維持します
- 異なる回転角のエンクロージャー内のTiO2-水ナノ流体の安定性と自然対流に関する実験的研究
- 小型化された伸縮性のある高レートのリニアスーパーキャパシタ
- ボロフェンの安定性とSTM画像に関する第一原理研究
- マウスにおける二酸化チタンナノ粒子の潜在的な肝臓、脳、および胚の毒性
- 光触媒活性と安定性を強化するためのIn2O3ナノ粒子ハイブリッド双晶六角形ディスクZnOヘテロ構造の水熱合成
- 階層的多孔質シリカライト-1カプセル化AgNPの調製と4-ニトロフェノール還元のためのその触媒性能