


復元されたmicroRNA-326-5pは、脳虚血/再灌流傷害におけるSTAT3の抑制を介して、ニューロンのアポトーシスを阻害し、ミトコンドリアの損傷を軽減します
要約
研究は、脳虚血/再灌流傷害(CI / RI)におけるマイクロRNA(miRNA)の役割を大いに調査しました。しかし、CI / RIにおけるmiR-326-5pの特定のメカニズムはまだとらえどころのないです。したがって、この研究は、CI / RIにおけるmiR-326-5p /シグナル伝達兼転写活性化因子-3(STAT3)軸のメカニズムを明らかにすることでした。 2つのモデル(初代ラット皮質ニューロンの酸素およびグルコース欠乏[OGD]とSprague-Dawleyラットの中大脳動脈閉塞[MCAO])は、それぞれinvitroおよびinvivoでCI / RIを模倣するために確立されました。機能喪失および機能獲得アッセイは、OGD処理ニューロンおよびMCAOラットを使用して実行されました。その後、OGD処理ニューロンの生存率、アポトーシス、酸化ストレス、ミトコンドリア膜電位、およびMCAOラットの脳組織の病理学的変化、アポトーシス、ミトコンドリア膜電位をテストしました。ミトフシン-2(Mfn2)、miR-326-5p、およびSTAT3の発現は、OGD処理ニューロンおよびMCAOラットの脳組織で検出されました。 Mfn2とmiR-326-5pは減少し、STAT3はMCAOラットのOGD処理ニューロンと脳組織で上昇しました。 miR-326-5pは、STAT3の発現を標的とし、負に調節されています。 miR-326-5pを復元するか、STAT3を減らすと、生存率が強化され、アポトーシスと酸化ストレスが抑制され、ミトコンドリア膜電位が上昇し、OGD処理ニューロンでMfn2の発現が増加しました。 miR-326-5pをアップレギュレートするか、STAT3をダウンレギュレートすると、病理学的変化が緩和され、MCAOを有するラットの脳組織におけるアポトーシスとミトコンドリア膜電位およびMfn2発現の上昇が抑制されました。この研究は、アップレギュレーションされたmiR-326-5pまたはダウンレギュレーションされたSTAT3が、Mfn2の発現を上昇させることによってCI / RIから保護することを明らかにしています。
はじめに
脳虚血/再灌流傷害(CI / RI)は、虚血性脳卒中が続く一種の脳損傷です[1]。大脳I / Rの最も特徴的な特徴は、再灌流後の初期の一過性脳虚血です[2]。 CI / RIはニューロンのアポトーシスを活性化し、海馬および皮質の損傷を引き起こします[3]。現在、最も一般的な臨床応用はCI / RIの血栓溶解療法です[4]。しかし、再灌流は活性酸素種(ROS)の生成を増加させ、細胞内DNA損傷、酸化ストレス関連損傷、タンパク質酸化、脂質過酸化を引き起こし、血液脳関門と浮腫をさらに悪化させます[5]。したがって、新しいターゲットオプションを検索する緊急性は、CI / RIの優先順位の最優先事項です。
マイクロRNA(miRNA)は、CI / RIにおける潜在的および予後的役割として広く考えられています[6、7]。たとえば、ある記事では、miR-202-5pが、虚血性損傷の中大脳動脈閉塞(MCAO)モデルラットにおける神経損傷と神経障害、および酸素とグルコースの欠乏(OGD)による細胞損傷を軽減することが示されています[8]。 。さらに、miR-98のアップレギュレーションは、I / R脳卒中のマウスの神経学的転帰を改善し[9]、miR-451の過剰発現はCI / RIのマウスの虚血性脳アポトーシスを軽減することがわかっています[10]。具体的には、miR-326-5pは内皮前駆細胞に対して血管新生促進能を持ち、急性心筋梗塞後の心機能を改善する可能性があります[11]。ただし、CI / RIでmiR-326-5pを中心とした研究はまだ始まったばかりです。シグナル伝達物質と転写活性化因子(STAT)は、CI / RIによって活性化されるユニークなタンパク質ファミリーです[12]。 STAT3は、バイオインフォマティクスのWebサイトでmiR-326-5pのターゲットとして予測されました。したがって、CI / RIにおけるmiR-326-5pを介したSTAT3の役割を研究しました。 STAT3は、炎症誘発性メディエーターを放出することにより、虚血性神経炎症プロセスと二次脳損傷を悪化させます[13、14]。具体的には、Janus活性化キナーゼ2(JAK2)/ STAT3経路は、CI / RIによって誘導される細胞アポトーシスをブロックする機能的な役割を果たします[15]。ミトフシン-2(Mfn2)はミトコンドリア融合因子であり、心脳血管I / RI [16]から保護し、虚血性脳損傷における低酸素誘発性ニューロンアポトーシスを改善する可能性があります[17]。 Mfn2がCI / RIに対して保護効果を発揮することも記録されています[18]。
一言で言えば、それほど詳細ではない調査により、CI / RIにおけるmiR-326-5pとSTAT3の複合的な役割が発見されました。そのため、この研究は、miR-326-5pがSTAT3を標的とすることでCI / RIを弱めるという仮説を立てて開始されました。
材料と方法
倫理声明
実験は揚州大学付属病院の動物管理使用委員会によって承認されました。揚州大学、そしてそれは揚州大学付属病院の実験用ラットの世話と使用へのガイドで実施されました。揚州大学。
実験動物
揚州大学(中国、揚州)の比較医学センターのオスの成体Sprague–Dawleyラット(6〜8週齢、250±30 g)は、特定病原体除去環境で飼育され、餌と水を自由に摂取できました。 (24±1)°C、(50±5)%湿度、12時間の明/暗サイクル。
セルの分離と識別
ラットを断頭により安楽死させ、脳を1mmに切断した 3 0.4%トリパンブルー溶液と9:1で混合した細胞懸濁液を構成し(トリパンブルーの最終濃度は0.04%)、血球計算盤で細胞密度と生存率を計算しました。セル(1×10 6 細胞/ mL)を、5×10 6 のL-ポリリジン(0.1 mg / mL)でプレコートした細胞培養フラスコに播種しました。 フラスコあたりの細胞。細胞が壁に付着したら、更新した培地(NeurobasalA + B27 + L-グルタミン)とインキュベートし、培地を2〜3日ごとに半分に交換して6〜7日間継続的に培養しました。
細胞の同定:ラット皮質ニューロンを利用してニューロンスライドを調製し、一次ウサギ抗ラットネスチン(NES)ポリクローナル抗体(1:200)、およびフルオレセインイソチオシアネート(FITC)標識ヤギ抗ウサギ免疫グロブリンとインキュベートしました。 G(1:50、両方ともCell Signaling Technology、マサチューセッツ州ビバリー、米国)。その後、細胞を抗蛍光消光剤で密封し、蛍光顕微鏡で観察しました。
OGDモデルの確立
ニューロンは、グルコースを含まないアールの溶液で培養され、95%N 2 の混合ガスにさらされました。 および5%CO 2 。陽圧換気の30分後、細胞が培養された低酸素環境が形成されました(invitroでのOGDによる虚血シミュレーション)。 90分間のインキュベーション後、グルコースを含まないアールの溶液を除去した後、細胞を維持細胞培地でインキュベートしました。細胞は、その後の実験(インビトロでの酸素-グルコース再灌流による再灌流シミュレーション)のために日常的に培養された。正常酸素下の通常の培地が対照として機能した。
ニューロンは対照群に分けられました。 OGDグループ、ネガティブコントロール(NC)グループ(スクランブルオリゴヌクレオチドでトランスフェクトされたOGD処理ニューロン)、miR-326-5pアゴミールグループ(miR-326-5pアゴミールでトランスフェクトされたOGD処理ニューロン)、sh-STAT3グループ(STAT3 shRNAでトランスフェクトされたOGD処理ニューロン)およびmiR-326-5pアゴミール+過剰発現(oe)-STAT3グループ(miR-326-5pアゴミールおよびpcDNA-STAT3ベクターでトランスフェクトされたOGD処理ニューロン)。オリゴヌクレオチドとベクターはすべてGenePharma(上海、中国)から提供されました。
リポフェクタミン2000(Invitrogen; Thermo Fisher Scientific)を使用して、スクランブルオリゴヌクレオチド、miR-326-5pアゴミール、STAT3 shRNAまたはmiR-326-5pアゴミール、およびpcDNA-STAT3を初代ラットニューロンにトランスフェクトし、トランスフェクション効率を逆転写定量でテストしました。 48時間後のポリメラーゼ連鎖反応(RT-qPCR)またはウエスタンブロット。
3-(4,5-ジメチルチアゾール-2-イル)-2,5-ジフェニルテトラゾリウムブロミド(MTT)アッセイ
ニューロンは1×10 6 で懸濁液にされました 細胞/ mLおよびそれぞれ0、24および48時間培養。無菌環境で、ニューロンを10%MTT溶液中で2〜4時間インキュベートし、結晶が十分に溶解するまで150μLのフォルマンザン溶解溶液(ジメチルスルホキシド)を添加しました。光学密度(OD)値は、マイクロプレートリーダーで570nmで測定されました。
フローサイトメトリーアポトーシスアッセイ
ニューロンを0.25%トリプシンで分離し、2000 rpmで遠心分離し、PBSに再懸濁しました。次に、ニューロンを2000 rpmで遠心分離し、ペレットを結合バッファーで再懸濁し、5μLのアネキシンV-FITCおよび5μLのヨウ化プロピジウム(PI)と連続してインキュベートしました。アポトーシス率を検出するためにフローサイトメーターを適用しました。
JC-1ミトコンドリア膜電位の検出
6ウェルプレートの細胞をPBSでリンスし、1mLの細胞培養液と1mLのJC-1染色作業溶液(50μLのJC-1、8mLの超純水と2mLのJC-1染色バッファー)(SolarbioScience 、北京、中国)および15分間インキュベートした。 1×JC-1染色バッファーは、4mLの蒸留水と1mLのJC-1染色バッファーで構成されました。インキュベーション後、細胞をトリプシン処理し、JC-1バッファーに再懸濁し、フローサイトメーターでテストしました。緑色蛍光の相対比率を計算しました。
酸化ストレスの検出
細胞内ROSレベルは、2,7-ジクロロフルオレセインジアセテート(DCF-DA)検出キット(Abcam)によって測定されました。要するに、ニューロンは0.25%トリプシンで分離され、再懸濁され、10μmのDCF-DAで30分間インキュベートされました。その後、495 nm / 529 nmの励起/発光を伴うDCF蛍光を、蛍光分光法(BD Biosciences)で測定しました。
細胞内malondialchehyche(MDA)、グルタチオン(GSH)含有量、およびスーパーオキシドジスムターゼ(SOD)活性の検出は、市販のアッセイキット(Beyotime Biotechnology Co.、Shanghai、China)を使用した化学測色法によって検出されました。 OD値はマイクロプレートリーダーで測定しました。
ラットモデルの確立
ペントバルビタールナトリウム(50μg/ kg)の腹腔内注射によりラットを麻酔した。首に正中切開を行い、外頸動脈を結紮し、総頸動脈を動脈クリップで塞いだ。 3–0シリコンコーティングされたモノフィラメントナイロン縫合糸を、光が抵抗するまで内頸動脈に注意深く挿入しました。一過性脳虚血の90分後、縫合糸を抜いて、さらに24時間再灌流を行った。偽のグループは、縫合糸が内頸動脈に入らなかったことを除いて、同じ治療を受けた。手術中、サーモスタットベッドを使用して直腸温を37±0.5°Cに維持しました[19]。
ラットを6つのグループ(n =6)に割り当て、偽のグループ、MCAOグループ(モデル化されたラット)、NCグループ(外側の脳室注射を行ったモデル化されたラット)を含むMCAOモデルを確立する前に、プラスミドまたはmiRを外側脳室に注射しました。 5μLのスクランブルオリゴヌクレオチド[100μM])、miR-326-5pアゴミールグループ(5μLのmiR-326-5pアゴミール[100μM]の側方脳室内注射を伴うモデル化ラット)、sh-STAT3グループ(モデル化されたラット5μLのSTAT3干渉ベクター[100μM]の脳室外側注射)およびmiR-326-5pアゴミール+ oe-STAT3グループ(5μLのmiR-326-5pアゴミール[100μM]および5μLの脳室外側注射を行ったモデルラットSTAT3過剰発現ベクター[100μM])[20、21]。
標本コレクション
ラットを麻酔で安楽死させ、心臓を灌流し、脳全体を採取した。脳損傷組織を1×1×1mm 3 に切断しました。 パラフィン切片の調製、RT-qPCR、ウエスタンブロット分析、ミトコンドリア膜電位検出のための組織塊。
大脳皮質におけるミトコンドリア膜電位の検出
SDラットの大脳皮質を分離し、1 mm 3 に切断しました。 。組織ブロックを300メッシュのナイロンネット上に置き、PBSを加えた。次に、細胞懸濁液を1000r /分で遠心分離し、70%エタノールを加え、再び1000r /分で遠心分離した。その後、サンプルを1 mLのPBSに再懸濁し、懸濁液(1×10 6 細胞(100μL)をRhodamine123色素溶液(10μL、5μg/ mL)と反応させ、フローサイトメーターで488nmで分析しました。
HE染色
ラットの脳組織を4%パラホルムアルデヒドに24時間浸漬し、パラフィン切片を調製し、4μmに切断し、40°Cの水に広げ、60°Cで焼きました。定期的に脱ロウおよび水和し、パラフィン切片をHE染色液で染色しました。 HE染色後、損傷したニューロンは、核の収縮、細胞浮腫、空胞化、および核の黒ずみを示した。光学顕微鏡(ニコン、日本)を使用して、画像と生存ニューロンの数(mm 2 )を取得しました。 )虚血性皮質の数を数えた。
TUNEL染色
パラフィン切片を脱ロウし、100%、95%、80%、70%のアルコールで脱水しました。次に、切片を4%パラホルムアルデヒドに浸し、0.1%Triton X-100クエン酸ナトリウムバッファーで20分間、TUNEL反応混合物で1〜1.5時間インキュベートしました。次に、切片をペルオキシダーゼ溶液とジアミノベンジジン(DAB)で現像し、ヘマトキシリンで対比染色し、勾配アルコールで脱水し、透過処理し、中性ガムで密封しました。切片を光学顕微鏡で観察して、TUNEL陽性細胞を数えた。核内の茶色がかった黄色の粒子は、TUNEL陽性細胞(アポトーシス細胞)として定義されました。
免疫組織化学
切片を脱ロウし、水和し、クエン酸抗原によって回収し、内因性カタラーゼを3%H 2 でブロックしました。 O 2 。各切片にヤギ血清(50μL)を加え、一次抗体(50μL)とインキュベートし、二次抗体(50μL)および西洋ワサビペルオキシダーゼ標識ストレプトアビジン(50μL)と反応させました。切片はDABによって現像され、ヘマトキシリンによって対比染色された後、勾配アルコール脱水、キシレン透過処理、および中性ガムシールが行われました。切片を顕微鏡で観察した(虚血性脳組織の細胞質または核は黄色または茶色がかった黄色の粒子であった)。 OD値が測定され、平均化されました。
RT-qPCR
組織および細胞からのトータルRNA抽出は、Trizol(Invitrogen)によって実行されました。 mRNA発現は、SYBR Green PCR Mix(Applied Biosystems、CA、USA)およびQuantStudio TM 6FlexリアルタイムPCRシステム(Applied Biosystems)を使用したqPCRによって分析されました。特定のステムループ逆転写プライマーおよびフォワード/リバースプライマー(BioTNT、上海、中国)は、miRNAレベルを分析するために設計されました。表1にすべてのプライマーを示します。 2 −△△CT miRNAまたはmRNAレベルの計算にはこの方法が採用されました。
<図>ウエスタンブロット分析
タンパク質サンプルを10%ドデシル硫酸ナトリウムポリアクリルアミドゲル電気泳動で分離し、ポリフッ化ビニリデン(PVDF)メンブレンにエレクトロブロッティングしました。その後、PVDFメンブレンをトリス含有生理食塩水中の5%脱脂粉乳でブロックし、一次抗体でプローブし、二次抗体で2時間再プローブしました。次に、タンパク質バンドを、ニトロブルーテトラゾリウムクロリドおよび5-ブロモ-4-クロロ-3-インドリルホスフェートを含むアルカリホスファターゼ緩衝液で分析し、ImageJソフトウェア(NIH、ベセスダ、メリーランド州、米国)によって定量化した。主な抗体は、STAT3(1:1000)、Mfn2(1:1000、米国マサチューセッツ州アブカム)およびグリセルアルデヒド-3-リン酸デヒドロゲナーゼ(GAPDH)(1:1000、Cell Signaling Technology、米国マサチューセッツ州ビバリー)でした。
デュアルルシフェラーゼレポーター遺伝子アッセイ
miR-326-5p野生型(WT)または変異体(Mut)結合部位を含むSTAT3 3'UTRを、STAT3-WTおよびSTAT3-Mutという名前のpmirGLOベクター(Promega、WI、USA)にクローニングしました。細胞を24ウェルプレートに2×10 4 で播種しました 細胞/ウェルおよび100ngのSTAT3-WTまたはSTAT3-Mutルシフェラーゼベクターおよび50nMmiR-326-5pアゴミールまたはそれアゴミールNCを70%コンフルエンスでコトランスフェクトしました。ルシフェラーゼ活性は、Dual-Luciferase Reporter Assay System(Promega Corporation、WI、USA)を使用して測定しました。
統計分析
SPSS 21.0(IBM、NY、USA)統計ソフトウェアをデータ分析に使用しました。データは平均±標準偏差(SD)として表されました。 2つのグループ間の差異は、スチューデントのt検定によって分析され、多重比較は、一元配置分散分析(ANOVA)と、それに続くTukeyの事後検定によって分析されました。 P 両面テストを表し、 P <0.05は統計的に有意であると見なされました。
結果
miR-326-5pのアップレギュレーションまたはSTAT3のダウンレギュレーションは、OGDで処理された皮質ニューロンのアポトーシスを減衰させます
1週間培養すると、ラットの皮質ニューロンが分化して成熟しました。顕微鏡的には、ニューロンは、より大きなニューロン体、より透明な細胞質、低密度の大きな核、および強い屈折率を示した。ニューロン体は円錐形または円形であり、樹状突起は千鳥状になって壁に付着していました(図1a、b)。 NESは、大脳ニューロンの特異的な化学的マーカーであり、免疫蛍光抗体法によって特異性の高いニューロンを特定します。ニューロンはNESに特異的に結合し、緑色の蛍光を示しました(図1c)。
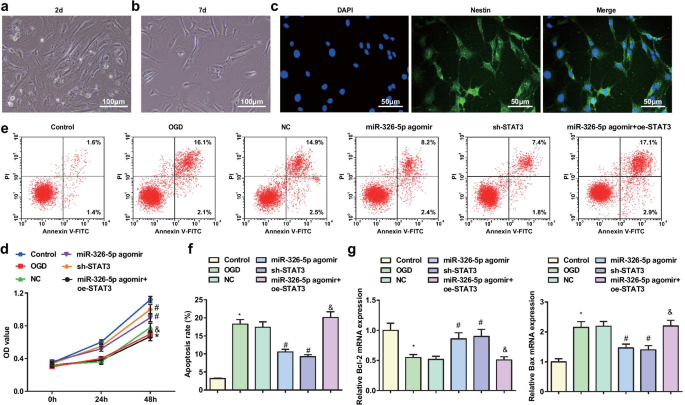
miR-326-5pのアップレギュレーションまたはSTAT3のダウンレギュレーションは、OGDで処理された皮質ニューロンのアポトーシスを減衰させます。 a 2次元付着培養を伴う脳ニューロン(スケールバー:100μm); b 7日間の付着培養を伴う脳ニューロン(スケールバー:100μm); c NES免疫蛍光抗体法で染色された7日間の付着培養を伴う脳ニューロン(スケールバー:50μm); d MTTアッセイによるOGD処理皮質ニューロンの生存率; e フローサイトメトリーによって検出されたOGD処理皮質ニューロンのアポトーシス; f OGD処理皮質ニューロンのアポトーシス率; g OGD処理皮質ニューロンにおけるBcl-2およびBaxmRNAの発現。 * P 対照群と比較して<0.05; # P NCグループと比較して<0.05; & P
MTTアッセイは、皮質ニューロンの生存能力を検出するために適用されました(図1d)。結果は、ラット皮質ニューロンの生存率が、OGDグループおよびmiR-326-5pアゴミール+ oe-STAT3グループと対照グループおよびmiR-326-5pアゴミールグループで損なわれたが、miR-326-5pでは増強されたことを示したアゴミールグループとsh-STAT3グループ対NCグループ(すべて P <0.05)。
ラット皮質ニューロンのアポトーシスはフローサイトメトリーによって決定され、Bcl-2およびBax mRNAの発現はRT-qPCRによって決定されました(図1e–g)。結果は、対照群およびmiR-326-5pアゴミール群と比較して、OGD群およびmiR-326-5pアゴミール+ oe-STAT3群は、アポトーシス率の増加、Bcl-2発現の減少、およびBax発現の増加を示したことを説明しました。 (すべての P <0.05)。対照的に、NCグループでは、miR-326-5pアゴミールグループとsh-STAT3グループ(すべて P )で、アポトーシス率が低下し、Bcl-2レベルが上昇し、Baxレベルが抑制されました。 <0.05)。
miR-326-5pのアップレギュレーションまたはSTAT3のダウンレギュレーションは、OGD損傷後の皮質ニューロンのMfn2およびミトコンドリア膜電位レベルを増加させます
OGDで処理された皮質ニューロンにおけるMfn2の発現は、ウエスタンブロット分析によって検出されました(図2a、b)。対照群およびmiR-326-5pアゴミール群に関しては、Mfn2タンパク質発現はOGD群およびmiR-326-5pアゴミール+ oe-STAT3群で減少したが、miR-326-5pアゴミール群およびshでは増加した。 -STAT3グループとNCグループ(すべて P <0.05)。
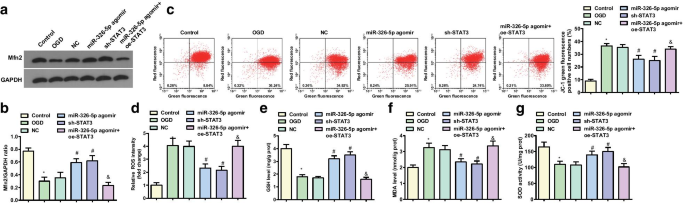
miR-326-5pのアップレギュレーションまたはSTAT3のダウンレギュレーションは、OGDで処理された皮質ニューロンのMfn2およびミトコンドリア膜電位レベルを増加させます。 a 、 b OGD処理皮質ニューロンにおけるMfn2タンパク質の発現; c OGD処理皮質ニューロンにおけるミトコンドリア膜電位レベルのJC-1フローサイトメトリー。 d miR-326-5pをアップレギュレートまたはSTAT3をダウンレギュレートした後のOGD処理皮質ニューロンのROS含有量。 e miR-326-5pをアップレギュレートまたはSTAT3をダウンレギュレートした後のOGD処理皮質ニューロンのGSH活性。 f miR-326-5pをアップレギュレートまたはSTAT3をダウンレギュレートした後のOGD処理皮質ニューロンのMDA含有量。 g miR-326-5pをアップレギュレートまたはSTAT3をダウンレギュレートした後のニューロンのSOD活性。 * P 対照群と比較して<0.05; # P NCグループと比較して<0.05; & P
JC-1によって検出されたミトコンドリア膜電位は、コントロールグループとmiR-326-5pアゴミールグループに関して、ミトコンドリア膜電位レベルがOGDグループとmiR-326-5pアゴミールで減少したことを発見しました(図2c、d)。 + oe-STAT3グループ(すべて P <0.05)。 NCグループと比較して、ミトコンドリア膜電位レベルは、miR-326-5pアゴミールグループとsh-STAT3グループ(すべて P )で増加しました。 <0.05)。
ROSとMDAの内容、およびOGDで処理された皮質ニューロンのGSHとSODの活動が決定され、結果は、対照群とmiR-326-5pアゴミールグループ、OGDグループとmiR-326-5pアゴミール+ oeに対して-STAT3グループは、ROSおよびMDAの含有量の増加と、GSHおよびSODの活動の障害を示しました(すべて P <0.05)。 NCグループと比較して、miR-326-5pアゴミールグループとsh-STAT3グループは、ROSとMDAの含有量が減少し、GSHとSODの活動が強化されました(すべて P <0.05)(図2e–g)。
miR-326-5pのアップレギュレーションまたはSTAT3のダウンレギュレーションは、皮質ニューロンの病理学的損傷を抑制し、CI / RIのラットのアポトーシスを抑制します
>HE染色は脳組織の病理学的損傷を観察するために採用され(図3a、b)、偽のグループでは、ニューロンは構造が正常であり、淡い赤色の細胞質、青い核、透明な核小体できれいに配置されていることを示しています。 MCAOグループ、NCグループおよびmiR-326-5pアゴミール+ oe-STAT3グループでは、壊死を除いて、ニューロンが無秩序に配置され、核膜の破裂、細胞構造の消失、核濃縮、深部を伴う暗く染色されることが観察されました。核の染色と大量の溶解。 miR-326-5pアゴミールグループとsh-STAT3グループでは、ニューロンの腫れが緩和され、ニューロンが整然と配置され、壊死細胞が減少し、MCAOグループとNCグループとの関係でより良い状態を示しています。
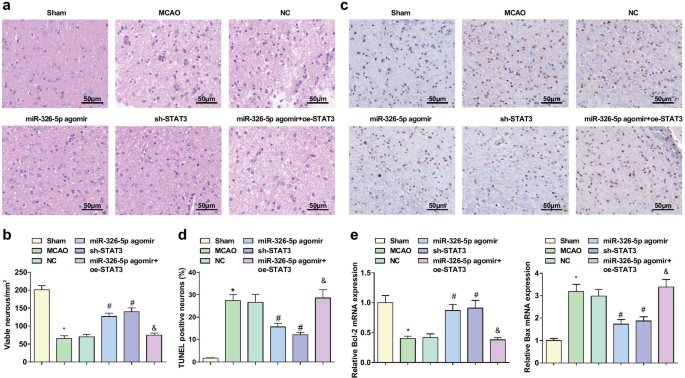
miR-326-5pのアップレギュレーションまたはSTAT3のダウンレギュレーションは、皮質ニューロンの病理学的損傷を妨げ、CI / RIのラットのアポトーシスを阻害します。 a HE染色で観察されたMCAOラットの脳組織の病理学的損傷(スケールバー:50μm); b miR-326-5pをアップレギュレートまたはSTAT3をダウンレギュレートした後のMCAOラット脳組織の無傷のニューロンの数。 c TUNEL染色によって検出されたMCAOラット脳組織の神経細胞アポトーシス(スケールバー:50μm); d miR-326-5pをアップレギュレートまたはSTAT3をダウンレギュレートした後のMCAOラット脳組織におけるTUNEL陽性率。 e miR-326-5pをアップレギュレートまたはSTAT3をダウンレギュレートした後のMCAOラット脳組織におけるBcl-2およびBaxmRNAの発現。 * P 偽のグループと比較して<0.05; # P NCグループと比較して<0.05; & P
TUNEL染色は、脳組織におけるBcl-2およびBaxmRNA発現のニューロンアポトーシスおよびRT-qPCRの検出に利用されました。結果は、(図3c–e)偽のグループおよびmiR-326-5pアゴミールグループと比較して、MCAOグループでTUNEL陽性率が増加し、Bcl-2 mRNA発現が減少し、BaxmRNA発現が増加したことを示しました。 miR-326-5pアゴミール+ oe-STAT3グループ(すべて P <0.05)。 miR-326-5pアゴミールとsh-STAT3グループではNCグループよりもTUNEL陽性率とBaxmRNA発現が低く、Bcl-2 mRNA発現が高いことが示されました(すべて P <0.05)。
miR-326-5pのアップレギュレーションまたはSTAT3のダウンレギュレーションは、CI / RIのラットの脳組織におけるミトコンドリア膜電位レベルを増加させます
ラット脳組織におけるMfn2タンパク質発現の検出は免疫組織化学によって行われ、その結果は、(図4a、b)偽グループおよびmiR-326-5pアゴミールグループとMCAOグループおよびmiR-326-5pアゴミール+ oe-STAT3グループ、より高いMfn2タンパク質発現は、NCグループの代わりにmiR-326-5pアゴミールグループとsh-STAT3グループで測定されました(すべて P <0.05)。
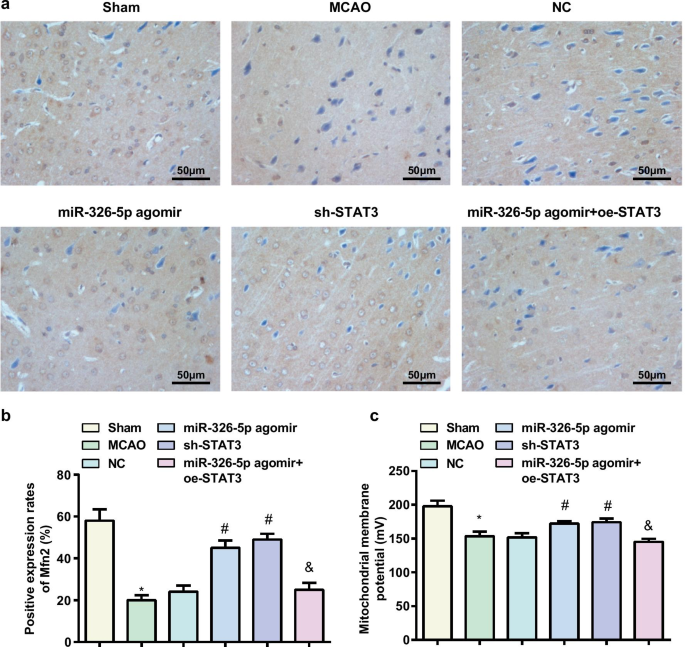
miR-326-5pのアップレギュレーションまたはSTAT3のダウンレギュレーションは、CI / RIのラットの脳組織におけるミトコンドリア膜電位レベルを増加させます。 a ラット脳組織における免疫組織化学によって検出されたMfn2発現(スケールバー:50μm); b miR-326-5pをアップレギュレートまたはSTAT3をダウンレギュレートした後のMCAOラット脳組織におけるMfn2陽性細胞の数。 c miR-326-5pをアップレギュレートまたはSTAT3をダウンレギュレートした後のMCAOラット脳組織のニューロンにおけるミトコンドリア膜電位。 * P 偽のグループと比較して<0.05; # P NCグループと比較して<0.05; & P
ミトコンドリア膜電位レベルの検出により、(図4c)偽グループおよびmiR-326-5pアゴミールグループと比較して、MCAOグループおよびmiR-326-5pアゴミール+ oe-STAT3でミトコンドリア膜電位レベルが低下することがわかりました。 NCグループとは対照的に、miR-326-5pアゴミールグループとsh-STAT3グループで増加しました(すべて P <0.05)。
miR-326-5pターゲットSTAT3
皮質ニューロンおよび脳組織におけるmiR-326-5pおよびSTAT3の発現は、RT-qPCRおよびウエスタンブロット分析によって検出されました。インビトロの皮質ニューロンでは、対照群(両方の P )と比較して、OGD群におけるmiR-326-5p発現の減少およびSTAT3発現の増加が示唆された。 <0.05)。 NCグループに関しては、miR-326-5pアゴミールグループ( P の両方)でmiR-326-5pの発現が上昇し、STAT3の発現が低下しました。 <0.05); STAT3の発現が減少しました( P <0.05)sh-STAT3グループ。 miR-326-5pアゴミールグループとは対照的に、STAT3発現はmiR-326-5pアゴミール+ oe-STAT3グループ( P )で上昇しました。 <0.05)(図5a–c)。
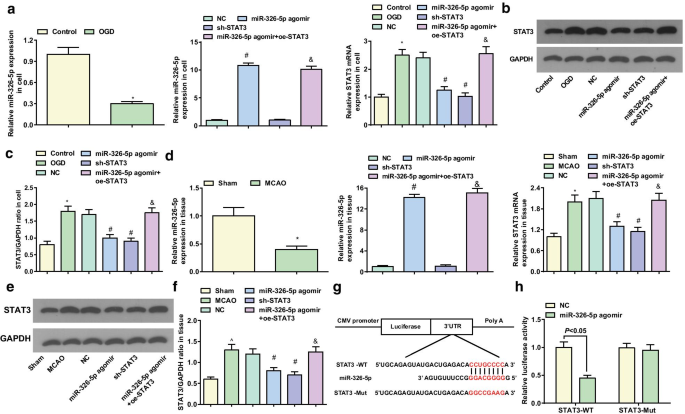
miR-326-5pはSTAT3を標的としています。 a invitroでのOGD処理皮質ニューロンにおけるmiR-326-5pおよびSTAT3mRNA発現。 b 、 c invitroでのOGD処理皮質ニューロンにおけるSTAT3タンパク質発現; d MCAOラット脳組織におけるmiR-326-5pおよびSTAT3mRNAの発現。 e 、 f MCAOラット脳組織におけるSTAT3タンパク質の発現; g バイオインフォマティクスソフトウェアによるmiR-326-5pとSTAT3の予測される結合部位。 h デュアルルシフェラーゼレポーター遺伝子アッセイによって検証されたmiR-326-5pとSTAT3の間のターゲティング関係。図 a – c 、* P 対照群と比較して<0.05; # P NCグループと比較して<0.05; & P miR-326-5pアゴミールグループと比較して<0.05。図 d – f 、* P 偽のグループと比較して<0.05; # P NCグループと比較して<0.05; & P
インビボの脳組織では、偽のグループに関連して、より低いmiR-326-5pおよびより高いSTAT3がMCAOグループに現れた(両方の P <0.05)。 NCグループと比較して、miR-326-5pの発現は上昇しましたが、STAT3の発現はmiR-326-5pアゴミールグループで減少しました(両方 P <0.05)。 STAT3の発現はsh-STAT3グループで減少しました( P <0.05)。対照的に、miR-326-5pアゴミールグループでは、STAT3発現はmiR-326-5pアゴミール+ oe-STAT3グループで上昇しました( P <0.05)(図5d–f)。
STAT3 3'UTR上の潜在的なmiR-326-5pターゲティング部位は、DIANAおよびmiRDBソフトウェアによって発見されました(図5g)。 WTmiR-326-5p結合部位とMut結合部位を含むSTAT33'UTRを構築し、pmirGLOプラスミドに挿入しました。 miR-326-5pのSTAT33'UTRへの結合は、ルシフェラーゼレポーターアッセイによってさらに検証されました。ルシフェラーゼレポーターアッセイは、miR-326-5pアゴミールがSTAT3-WTの相対的なルシフェラーゼ活性を低下させたが、STAT3-Mutは低下させなかったことを明らかにし(図5h)、STAT3がmiR-326-5pの直接の標的遺伝子であることを示唆しています。
>ディスカッション
CI / RIは脳血管死の主な原因です[22]。 miRNAはCI / RIに関与していることが示されています[23]。ただし、miR-326-5p関連のメカニズムの包括的な理解はまだCI / RIでは不十分です。したがって、この研究は、miR-326-5pとSTAT3の逆数の組み合わせに重点を置いて、CI / RIにおけるmiR-326-5pの具体的な役割を明らかにすることを目的としています。生産的に、この研究は、miR-326-5pのアップレギュレーションがCI / RIを弱め、STAT3阻害を介してMfn2発現を上昇させることを宣言しました。
当初、miR-326-5pの発現はinvivoおよびinvitroでCI / RIが低下することが確認されました。続いて、一連のアッセイをアレンジし、miR-326-5pをアップレギュレートすると、ニューロンの生存率とミトコンドリア機能が強化され、Mfn2の発現が上昇し、酸化ストレスとアポトーシスが抑制されることがわかりました。さらに、ラットでのin vivo実験により、CI / RIで復元されたmiR-326-5pの機能的役割がさらに検証されました。実際のところ、高いmiR-326は、脳腫瘍の一般的なタイプである膠芽腫患者の全生存期間が長いことと相関していることが証明されています[24]。また、miR-326の発現はパーキンソン病を減少させることがさらに証明されており、miR-326はドーパミン作動性ニューロンのアポトーシスを抑制し、炎症反応を減少させる可能性があります[25]。それを除いて、miR-326-5pの減少は心筋梗塞の過程で内皮前駆細胞に現れ、過剰発現する内皮前駆細胞の導入は心機能の回復を促進する可能性があります[11]。 miR-326のアップレギュレーションが、アルツハイマー病のマウスの行動機能を改善し、ニューロンの生存能力を高め、ニューロンのアポトーシスを抑制することを示す研究があります[26]。さらに、miR-326ミミックによるmiR-326のアップレギュレーションは、パーキンソン病のドーパミン作動性ニューロンにおける誘導型一酸化窒素シンターゼを抑制できることが報告されています[27]。
この研究では、STAT3がmiR-326-5pの標的遺伝子であることがわかりました。高度な研究によって裏付けられており、STAT3の発現はヒト子宮内膜癌幹細胞においてmiR-326によって負に調節されていることが示されています[28]。本研究では、STAT3の発現がCI / RIで増強されることを測定しました。現在、I / R損傷とOGDがSTAT3発現を誘導して増加させることがわかっています[29]。興味深いことに、STAT3mRNAの発現はI / R動物で増加する傾向があります[30]。さらに、STAT3タンパク質の発現はCI / RIでアップレギュレートすることが報告されています[31]。次に、私たちの研究は、STAT3のダウンレギュレーションがCI / RIラットとOGD処理ニューロンに治療効果をもたらすことを明らかにしました。研究によって裏付けられているように、miR-31誘導は、虚血性脳卒中におけるJAK / STAT3経路の不活性化によって酸化ストレスを阻止すると結論付けられています[32]。また、JAK2 / STAT3経路の阻害は、CI / RIの酸化ストレス障害に有益です[31]。さらに、JAK / STAT3経路のノックダウンは、細胞の生存率を高め、虚血性脳損傷における酸化ストレスとニューロンのアポトーシスを減少させることが確認されています[33]。最近、阻害されたJAK / STAT3シグナル伝達経路が、心筋I / R損傷における心筋梗塞を緩和することが目撃されています[34]。示されているように、枯渇したSTAT3は、白質損傷のあるラットの神経細胞アポトーシスを弱体化させます[35]。さらに、ニューロンのアポトーシスの抑制、および脳梗塞のサイズの緩和は、CI / RIのラットにおけるJAK2 / STAT3阻害に起因します[36]。
最後に、Mfn2発現に対するmiR-326-5pとSTAT3の効果に焦点を当て、CI / RIおよびOGD処理ニューロンを有するラットのMfn2レベルの低下は、miR-326-5pを復元するか、STAT3をサイレンシングすることによって上昇する可能性があることを発見しました。 。以前の研究で示唆されているように、Mfn2は再灌流の後期に減少し、Mfn2の発現が不十分な場合、オートファゴソーム形成の抑制とオートファゴソームおよびリソソームの融合によってCI / RIが悪化することが報告されています[18]。さらに、Mfn2は、古い肝細胞のI / R損傷での消失さえも減少させることが示唆されています[37]。ただし、Mfn2に対するmiR-326-5pおよびSTAT3の調節メカニズムについて論じた研究はほとんどありません。
結論
一般に、この研究は、上昇したmiR-326-5pがニューロンのアポトーシスを阻害し、ニューロンの病理学的損傷を軽減し、CI / RIでのSTAT3のダウンレギュレーションを介してMfn2の発現を増加させるメカニズムを解明しました。この研究は、CI / RIにおけるmiR-326-5pおよびSTAT3の潜在的なメカニズムを更新する可能性があります。ただし、CI / RIの分子メカニズムをさらに発展させるには、さらに多くの研究が必要です。
データと資料の可用性
該当なし。
略語
- miRNA:
-
マイクロRNA
- STAT3:
-
シグナル伝達兼転写活性化因子-3
- OGD:
-
酸素とブドウ糖の欠乏
- MCAO:
-
中大脳動脈閉塞
- ROS:
-
活性酸素種
- STAT:
-
シグナル伝達物質と転写活性化因子
- Mfn2:
-
ミトフシン-2
- FITC:
-
フルオレセインイソチオシアネート
- NES:
-
ネスティン
- NC:
-
ネガティブコントロール
- Oe:
-
過剰発現
- RT-qPCR:
-
逆転写定量的ポリメラーゼ連鎖反応
- OD:
-
光学密度
- PI:
-
ヨウ化プロピジウム
- MDA:
-
Malondialchehyche
- GSH:
-
グルタチオン
- SOD:
-
スーパーオキシドジスムターゼ
- DAB:
-
ジアミノベンジジン
- PVDF:
-
ポリフッ化ビニリデン
- WT:
-
野生型
- ミュート:
-
ミュータント
- SD:
-
標準偏差
- ANOVA:
-
一元配置分散分析
ナノマテリアル
- 火災による損傷と予防
- microRNA-342-5pのダウンレギュレーションまたはWnt3aのアップレギュレーションは、血管新生を阻害し、アテローム性動脈硬化症マウスのアテローム性動脈硬化症のプラーク安定性を維持します
- ICAをロードしたmPEG-ICAナノ粒子の調製とLPS誘発性H9c2細胞損傷の治療におけるそれらの応用
- ゾル-ゲル法によるナノ構造シリカ/金-セルロース結合アミノ-POSSハイブリッド複合材料とその特性
- PEG化リポソームを介したブファリンの改善された抗腫瘍効果および薬物動態
- 蒸発誘起自己組織化と強化されたガス検知特性によるワームホールのようなメソポーラス酸化スズの容易な合成
- サイズ、修正、欠陥、およびドーピングによるグラフェン仕事関数の設計と調整:第一原理理論研究
- 酸化還元刺激によって誘発される放出のためのナノ粒子を標的とする新規の二重ミトコンドリアおよびCD44受容体
- アルギン酸ナトリウム電解質-スルホン化酸化グラフェン生体膜によるプロトン伝導性とメタノール透過性の低下の強化
- 水熱反応によりレモンジュースから製造された蛍光炭素量子ドットの材料と光学特性
- ゾルゲル法で合成されたPrドープペロブスカイトマンガン酸塩La0.67Ca0.33MnO3ナノ粒子の微細構造、磁気、および光学特性