


原子間力顕微鏡ナノスケールの機械的マッピングによる大腸菌遺伝子型の特徴的な高分子の同定
要約
微生物株の分類は、従来、分子法に基づいており、研究された細菌株の形態学的特徴はめったにありません。この研究では、AFM機械的マッピングを介して細菌表面の高分子構造を明らかにしました。その分解能は、ナノスケールの先端サイズだけでなく、標本の機械的特性によっても決定されました。この技術により、簡単な標本の準備と柔軟な作業環境で微生物株の膜構造のナノスケール研究が可能になり、電子顕微鏡法とラベル対応の生化学的分析方法における複数の制限を克服しました。細胞表面に位置する特徴的な高分子は、表面層タンパク質と見なされ、 Escherichia coli に特異的であることがわかりました。 遺伝子型。平均分子サイズは38〜66 nmの範囲の直径で特徴付けられ、分子形状は腎臓のようなものまたは円形でした。結論として、表面高分子構造は、 Eにリンクする独特の特性を持っています。コリ 遺伝子型。これは、細胞形態に対するゲノム効果がAFMメカニカルマッピングを使用して迅速に特定できることを示唆しています。
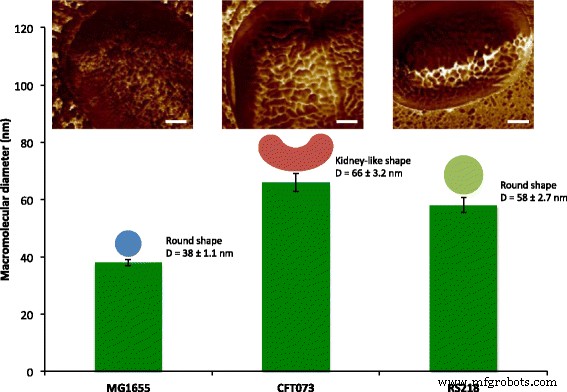
Eの表面高分子の定量化。コリ AFMメカニカルマッピングを使用したセル。 3つの Eの細胞表面の表面高分子。コリ 遺伝子型、MG1655、CFT073、およびRS218は、38〜66 nmの範囲のサイズで、円形または腎臓のような形状で特徴づけられました。地形画像は、スケールバー=200nmの接着マッピングで色付けされました。
背景
光学顕微鏡の解像度が低く、電子顕微鏡の作業環境が限られているため、微生物分野の研究者は、細菌細胞の外観を使用することをめったに検討せず、代わりに、ゲノム断片の同定、タンパク質の発現などに分子または化学分析法を採用します。当然のことながら、これらの方法には、労働集約的で時間がかかるなど、多くの欠点があります。したがって、より直接的で、効率的で、柔軟なアプローチが必要です。 1982年にBinnigらによって発明された原子間力顕微鏡(AFM)は、レーザービームで監視されるナノスケールのプローブを使用して、ナノスケールまたは原子分解能で試料の表面を観察するように設計されています[1]。物理的なプローブチップを介して形態を画像化するこの技術は、光学顕微鏡と電子顕微鏡の両方の分解能の限界と環境の制限を克服し、簡単な標本の準備や周囲の空気や流体の条件での柔軟な作業環境など、多くの利点があります[2、 3]。それでも、微生物研究に関して、AFMの現在のアプリケーションは、主に細菌細胞の静的または動的形態またはべん毛と線毛の発現を定性的に画像化することを含みます[4,5,6]が、微生物細胞と細胞特性の定量分析。
この研究では、 Escherichia coli の表面研究にAFMが選択されました。 ( E.coli )細胞、および個々の細菌の形状と寸法は、AFMトポグラフィーと位相画像によって観察されました。さらに、同時機械的マッピングにより、表面成分に関する追加の生体力学的情報が明らかになり、高分子と周囲のマトリックスとの間の接着特性のわずかな違いが、チップとサンプルの間の各物理的接触中に検出できました。このような高度な技術を微生物分野に適用して、3つの Eを検討しました。コリ 表面高分子の同定のために1つの実験室株と2つのヒト病原性株を含む遺伝子型。結果は、この技術が試料の機械的分布を感知することにより、AFMチップのスケールを超える画像解像度を提供できることを示しました。結論として、表面科学におけるそのような発展は、微生物分野で働く研究者に細胞の外観の詳細を提供するだけでなく、別のバイオまたはナノ材料システムの表面特性の知識にも貢献することを示唆します。
メソッド
微生物サンプル
3つの E。コリ この研究でテストされた菌株は、臨床的に分離され、国立成功大学分子医学研究所のチンハオテン教授の研究室によって提供されました。 MG1655は、 Eの腸および野生型の実験室株です。コリ 幼稚園から高校まで、および他の2つの菌株はヒト病原体です。尿路感染症の主な原因であるCFT073と、乳児の新生児髄膜炎に関連するRS218です[7]。
機能化された基板
固体表面と微生物細胞の間の共有結合のために、表面修飾の2つのステップが適用されました。最初に、3-アミノプロピルトリエトキシシラン(APTES、Sigma-Aldrich Co. LLC、米国)溶液を使用して、最初のNH 2 を形成しました。 -安定したAPTESコーティングが安定したSi-O結合に寄与した表面の官能化層-酸化された表面からAPTESとOによって提供されたSi。 2つのCOOH機能を持ち、グルタルアルデヒドはNH 2 に結合します 1つのCOOHでAPTESの機能を発揮し、NH 2 で機能します 他のCOOHとバクテリアの表面に。
きれいな基板をエタノール中の5%APTESの混合物を含むAPTES溶液に1時間浸し、エタノールとddH 2 ですすいだ。 O.スライドを窒素流を使用して乾燥させた後、グルタルアルデヒド溶液(PBS中2%)に一晩入れ、PBSで洗浄しました。
サンプル準備
Eの単一コロニー。コリ 菌株は溶原性ブロス(LB)寒天プレートから選択され、LBブロスで培養されました。 12時間の培養時間の後、細菌溶液を事前に平衡化した新鮮なLBブロスで1:100に希釈しました。微生物培養のためにさらに12時間後、細菌溶液を1500× g で遠心分離しました。 (4000 rpm)3分間、LBブロスに再懸濁し、このプロセスを2回繰り返します。 200マイクロリットルの細菌溶液を機能化された基質に滴下し、30分間静置しました。次に、標本を蒸留水に2回沈めて、付着していない細胞を取り除き、すぐに周囲空気中のAFMで画像化しました。
AFMの特性評価
微生物標本の表面検査には、AFM装置(Bruker Nano、サンタバーバラ、カリフォルニア州、米国)と、校正されたばね定数が0.7 N / m、先端半径が10nmの窒化ケイ素プローブを選択しました。 AFMスキャンレートとラインピクセルは、トポグラフィの最初の検出のスキャンサイズが10μmの場合、それぞれ0.5 Hzと256ラインでした。次に、パラメータは、スキャンサイズが2μmの場合、0.3Hzと512ラインに設定されました。詳細な観察。ナノメカニカルマッピングには、PeakForce定量的ナノメカニカル(QNM)モードが使用され、表面の接着特性は、引き抜き力-距離曲線の中で最大の引力から計算されました。
ストレプトコッカスミュータンスに関する以前の研究 は、細菌表面で2時間の機械的進化を示し、継続的なAFM機械的マッピングによって監視され、微生物サンプルはそのような期間内に生存していることが確認されました[8、9、10]。 Eの実行可能性を確保するため。コリ この作業で使用されたサンプル、細菌サンプルの事前調査が実施され、4時間の表面接着の継続的な変化は、サンプル準備後少なくとも4時間は細胞が生きていることを意味しました(追加ファイル1に示されています)。その結果、この作業でテストされた微生物サンプルは、標本の準備後2時間以内にAFMによって測定されました。すべての E。コリ 遺伝子型は個別に異なる時期に培養され、AFM測定は検体調製直後に実施されました。言い換えれば、細菌サンプルは検査を待つために並んでいなかったので、 E間の差に対する保持時間の影響。コリ 菌株は最小限に抑えられました。各 Eの定量的データ。コリ この研究で2時間以内に行われた測定からひずみが収集されました。
統計分析
この作業の統計分析には、Prism(GraphPad Software、USA)を使用しました。細胞の長さと高分子のサイズは、平均の標準誤差(SEM)とともに平均値として表されました。 E間の多重比較。コリ 遺伝子型は、通常の一元配置分散分析(ANOVA)を使用して処理されました。 95%の信頼水準( p <0.05)が選択され、アスタリスクは検出された有意差の程度を示します。各菌株のサンプル数nは> 40でした。
結果と考察
複数のマッピングによる表面の超微細構造
細菌サンプルで10μmの観察スケールでAFMを使用してスキャンする場合、いくつかの単一の E。コリ MG1655細胞を見ることができ、地形画像から3次元の細胞形状を観察することができました(図1a)。二次元で示された細胞の明確な輪郭は、たわみ誤差画像(図1b)によって得られ、細菌細胞のほかにいくつかの細管が見つかりました。波状のフィラメント(図1c)は、別の研究で報告された微生物のべん毛の外観と一致しており、べん毛などの発見が確認されました。また、短くて髪の毛のような線毛も見られました[11]。単一微生物細胞の詳細な研究のためにプローブの観察領域を減らすと、図1dに示すように、トポグラフィーは細胞表面間で垂直方向にわずかな違いを示し、たわみ誤差画像はより多くの形態学的情報を提供するように見えましたが、収集された環境ノイズは、細胞表面の超微細構造を調査するには多すぎました(図1e)。先端と被検物との接触時に試料の生体力学的特性を同時に測定したところ、接着力マッピングから明らかなように、細菌表面は実際には特定の形状とサイズの高分子で構成されていることがわかりました(図1f);したがって、地形およびたわみエラー画像の形態学的解像度は、生体力学的情報によってさらに改善されました。
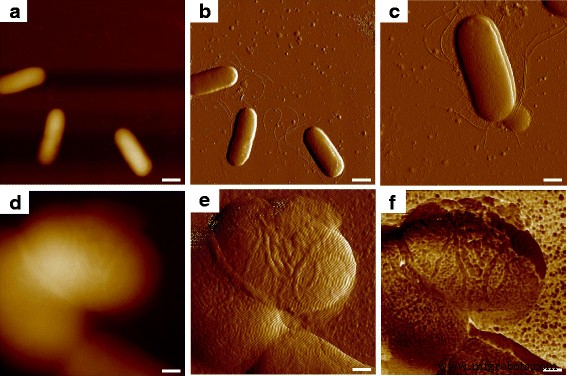
Eの表面超微細構造。コリ AFMマルチマッピングを使用するMG1655。 a および b 地形およびたわみ誤差の画像であり、 c べん毛と線毛を発現した細菌細胞の詳細なたわみ誤差画像でした。次に、単一のセルに焦点を合わせました。ここで、 d および e 地形およびたわみ誤差の画像であり、 f 対応する接着マッピングでした。スケールバー=1μmin a および b 、 c で500nm 、および d で200nm – f
図2aでは、 Eによるべん毛の表現。コリ MG1655がはっきりと見られ、フィラメントのサイズは図1bのものと同様であり、接着特性は基板よりも比較的低くなっています。さらに、細菌の表面は、周囲のマトリックスと比較した場合に接着性が低いことを特徴とする円形の成分で構成されていることがわかった。この観察結果は、マウスの皮膚の組織層に関する以前の発見と類似しており、繰り返し発生する同等の粒子は高分子であると見なされました。高分子の構造は、分子間領域で見られるものよりも密度が高く、一貫しているため、接着性能の違いがあります。簡単に感知できます[12]。グラム陰性菌の細胞外皮の最外層は、図2bに示すように、自己組織化タンパク質の層であり、表面層(S層)タンパク質として知られています[13]。 S層構造は従来、電子顕微鏡で測定されていましたが、真空環境と導電性コーティングの要件により、タンパク質に関するネイティブ情報とリアルタイム情報が失われていました。いくつかの研究はS層タンパク質を抽出し、AFMスキャンのためにそれらを雲母基板上に再構築しましたが、S層構造のinsituおよびリアルタイム性能の結果は欠けていました[14、15]。細胞構造と電子顕微鏡による微生物表面の以前の画像に基づいて、観察された高分子をS層タンパク質と見なしました[16]。
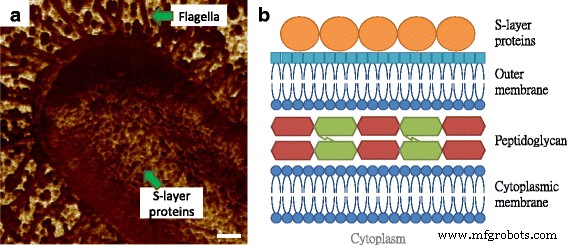
Eの垂直構造と表面構造の図。コリ 細胞。 a Eの表面高分子。コリ AFM接着マッピングによって画像化されたMG1655細胞。 b グラム陰性菌の細胞外皮の分子構造。これは、細胞質膜、ペプチドグリカン、外膜、およびS層で構成されています。スケールバー=200 nm
AFMと透過型電子顕微鏡(TEM)の測定値を比較すると、前者には後者に比べていくつかの利点があります。たとえば、標本の準備が簡単で、実験要件が制限されておらず、生物学に適したイメージングアプリケーションです。生物学分野でのTEMの選択は、細胞間小器官を見る能力と、得られる超高解像度(通常はナノメートルまたはサブナノメートル)のイメージングのために、一般的に選択されます。現在の作業でのAFM接着マッピングは、特徴の解像度を改善し、先端のサイズではなく、サンプル自体の固有の構造に依存する方法で表面高分子の配置を提示しました。さらに、このアプローチにより、数十マイクロメートル四方の領域全体で微生物表面のナノスケールの解像度が可能になります。
形態学的特徴のゲノム操作の違い
Eの表面微細構造を観察した後。コリ MG1655細胞、興味深い問題の1つは、高分子が他の菌株にどのように配置されているかです。ヒト病原体 E。コリ したがって、CFT073とRS218は、同じ実験パラメータを使用してAFMによって検査され、図3a〜cに示すように、10μmの特徴サイズでこれら3つの遺伝子型間で細胞の形状と寸法に有意差はありませんでした。接着力マッピングを使用してS層タンパク質を特定し、さまざまな形状とサイズの異なる構造が異なる E間で検出されました。コリ 図3d–fに示すように、ひずみ。比較を容易にするために、 Eの詳細な接着マッピング画像。コリ 菌株は図3g–iに表示されています。表面高分子は、MG1655およびRS218セルで丸い形状をしていると特徴づけられましたが、分子直径は異なり、38±1.1 nm( n )でした。 =80)MG1655および58±2.7 nm( n の場合 =46)RS218の場合。一方、CFT073細胞は、腎臓のような独特の形状のS層タンパク質を持ち、2つのエンドポイント間の長さの差は66±3.2 nm( n )でした。 =44)。多重比較のためにこれら3つの遺伝子型のS層タンパク質のサイズを分析した後、結果はこれらの菌株間の有意差を示しました(図4)。
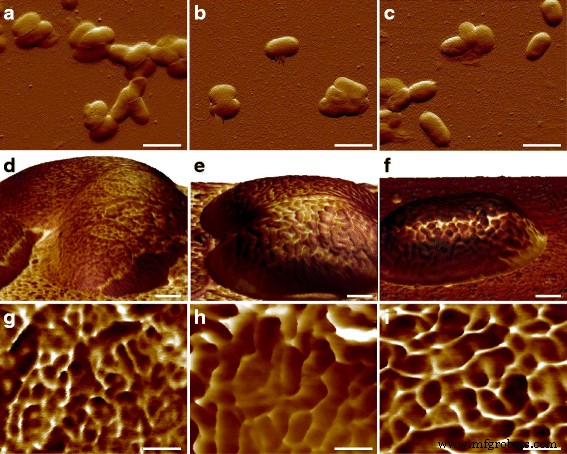
Eの形態的特徴。コリ 遺伝子型。上段は a のたわみ誤差画像を表示しています MG1655、 b CFT073、および c RS218。真ん中の行は、 d の接着マッピングで色付けされた3Dトポグラフィを示しています。 MG1655、 e CFT073、および f RS218セル。下の行は、 g の詳細な接着マッピングです。 MG1655、 h CFT073、および i RS218。接着マッピングでは、色が濃いほど接着性能が低く、その逆も同様です。 a のスケールバーは2μmでした – c 、 d の場合は200nm – f 、および g の場合は100nm – i
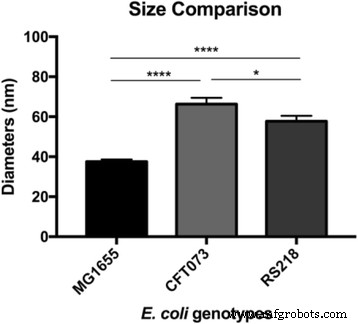
Eの分子サイズ。コリ 遺伝子型。表面タンパク質の直径はAFMによって検出され、多重比較のために一元配置分散分析によって処理されました。 **** p <0.001および* p <0.05
微生物のS層は、過酷な環境からの細胞の保護、食作用の攻撃、捕食性細菌など、多くの機能で重要な役割を果たすと報告されています。さらに、S層は効果的なコロニー形成を可能にするアドヘシンとしても機能します[17]。 S層構造は、TEMによって十分に研究されており、中心から中心までの間隔が4〜35nmのさまざまな格子タイプに分類されています[16]。私たちのAFM結果と文献からのTEMレポートの間の変化は、TEMがS層構造の2D形態を与え、AFMが細胞のラジアンと粗さによってもたらされる複数の影響を含む3Dトポグラフィーをキャプチャする異なるイメージング方法として考えられました。 AFMプローブの形状。
さまざまなタイプのS層構造は、元々、さまざまな分類学的特性を使用して細菌種を区別することが可能であると考えられていましたが、単一の種であっても、微生物の染みはタンパク質のさまざまな格子を持っている可能性があることがわかりました[13、16、18 19]。いくつかの研究はフィラメント形成におけるS層の役割を調査しましたが、細胞膜上のタンパク質の種類と E間のサイズのゲノム多様性。コリ 遺伝子型、S層タンパク質の違いはめったに注目されていません[20、21、22]。現在の研究の結果は、 E間の形態学的特徴の違いを明らかにした。コリ MG1655、CFT073、およびRS218は、表面高分子の外観がおそらく個々の Eに特異的であることを示唆しています。コリ 遺伝子型。
結論
この作業では、細菌表面に関するゲノム固有のナノ構造情報が、高分子と周囲のマトリックスとの間の接着の違いを区別するAFMメカニカルマッピングによって検出されました。グラム陰性菌の分子構造によれば、微生物細胞の表面高分子は表面層タンパク質と見なされていた。それらの高分子の配置およびサイズは、試験された Eに特異的であることが見出された。コリ 異なる形状とサイズの遺伝子型。これらの違いは統計分析によって有意であることが示されています。結論として、細菌のS層構造はゲノムに依存しており、微生物関連疾患または微生物株の迅速な診断のための潜在的な方法である可能性があると考えています。 S層特性の実用化を実現するためには、補体カタログにより多くの細菌遺伝子型の検討が必要です。現在、細菌の形態的特徴と生理学的/病理学的性能を結び付けるデータベースを確立しており、AFM検査の実用化に有望な進歩になると信じています。
略語
- AFM:
-
原子間力顕微鏡
- ANOVA:
-
一元配置分散分析
- APTES:
-
3-アミノプロピルトリエトキシシラン
- LB:
-
溶原性ブロス
- QNM:
-
定量的ナノメカニカル
- S層:
-
表面層
- TEM:
-
透過型電子顕微鏡
ナノマテリアル
- IBMの科学者がナノスケール用の温度計を発明
- カヴリ賞を受賞したAFMパイオニア
- ビッグオイルの次の噴出はナノスケールです
- 非導体はナノスケールで電流を伝導します
- Escherichia coliに対する酸化銅ナノ粒子の生体合成、特性評価、および抗菌力の評価
- ナノチャネルにおける油輸送に対する表面効果:分子動力学研究
- ナノスケール脂肪フラクタルからの小角散乱
- 偏位したトランケートされたAgナノプリズム二量体への局在表面プラズモン共鳴依存性
- 修飾された自由表面エレクトロスピニングを使用した高品質ナノファイバーのハイスループット製造
- AFMチップベースの動的耕起リソグラフィーを使用したポリマー薄膜上での高スループットのナノスケールピットの製造
- TiをドープしたMgAl2O4ナノリンの表面の欠陥