


食用菌根菌Tricholomacrassumから合成されたタンパク質でキャップされた金ナノ粒子の抗菌性、アポトーシス性、および癌細胞の遺伝子送達特性の評価
要約
追加のキャッピングステップなしで高機能タンパク質コートを備えた明確な幾何学的形状の金ナノ粒子の生合成はめったに報告されません。この研究では、食用菌根菌 Tricholoma crassum からのタンパク質コーティングされた金ナノ粒子のグリーン合成について説明します。 (バーク。)Sacc 。 ナノ粒子のサイズ範囲は5〜25 nmで、形状はさまざまでした。分光分析は、製造中のより長い反応期間での吸収極大の赤方偏移と、pHの増加での青方偏移を示しました。これらは、分光法、SEM、TEM、AFM、XRD、およびDLSで特徴づけられました。粒子サイズは、合成パラメータを変更することで変更できます。これらは、細菌、真菌、および多剤耐性病原菌に対して強力な抗菌活性を持っていました。これらはまた、細菌の増殖動態および真菌胞子の発芽に対して阻害効果を有していた。これらは、コメットアッセイでテストしたときに真核細胞でアポトーシス特性を示しました。さらに、粒子は、肉腫癌細胞に送達される遺伝子の付着部位として利用された天然の40kDaタンパク質で覆われています。本研究はまた、治療への応用のために、溶血アッセイを使用してこれらのナノ粒子の安全な投与量を最適化することを試みた。発酵槽でのナノ粒子の大規模生産および粒子の他の可能な用途が議論されています。
背景
ナノ粒子が農業、医薬品、家庭用品に不可避で広く適用されるようになると、有害な影響とそれを減らす方法を理解する必要性が高まります[1]。生物またはその酵素を使用したナノマテリアルのグリーン合成には、環境に優しく、費用効果が高く、治療用途に安全であるという利点があります[2]。微生物の中で、糸状菌は、より多くの酵素を分泌する能力があるため、ナノ粒子を合成する能力が高くなります[3、4]。細胞外金ナノ粒子(AuNP)は、いくつかの真菌を使用して生成されていますが[5、6、7]、これらのレポートでは、これらの粒子を覆う天然タンパク質のコートについてはほんのわずかしか言及されていません[8、9]。
医学では、多剤耐性菌の登場により、ナノ粒子は耐性を生じさせないため、代替の選択肢となります[10]。 AuNPは、腫瘍細胞の治療と診断、およびナノキャリアベースの遺伝子送達において特に有望です[11、12、13]。生理学的条件下では、AuNPは細胞膜を介して低い透過性を示しますが、腫瘍細胞では、浸透および保持(EPR)効果の強化により取り込みが強化されます[14]。この取り込みは、AuNPがタンパク質でキャップされている場合にさらに強化されます。プロテインキャップは、コロイド状態のナノ粒子を安定させ、送達用の薬物または遺伝子のドッキングサイトを提供するのに役立ちます[14]。ただし、キャッピングのプロセスには追加の手順が含まれます。天然タンパク質キャップの提供は、ほとんど危険な化学的経路の適用を排除します[5]。その重要性にもかかわらず、天然タンパク質コートを用いた貴金属ナノ粒子のグリーン合成に関する報告は限られています[15、16]。
治療にAuNPを適用する前に、分析すべき重要な側面は、ナノ材料と細胞膜との生体適合性です[6]。別の側面は細胞毒性であり、AuNPは赤血球を凝集させる可能性があります[13]。したがって、危険な側面に重点を置き、安全な範囲内で使用する必要があります[7、17]。
私たちの研究室は、これまでのところ、細胞外銀ナノ粒子を生成する唯一の食用菌根菌を報告しました。菌は Tricholoma crassum (バーク。)サック[2]。ここでは、 TからのAuNPのグリーン合成について説明します。 crassum サイズ範囲は5〜25 nmで、形状はさまざまです。これらは、細菌、真菌、および多剤耐性(MDR)病原性細菌に対する抗菌活性について特徴づけられ、分析されました。これらは、バクテリアの成長速度と真菌胞子の効力に抑制効果をもたらしました。最も重要なことは、粒子が自然にタンパク質でコーティングされていることです。これらの粒子を、癌細胞への遺伝子送達の媒体としての有効性についてテストしました。これらの粒子の生体適合性と毒性をチェックするために溶血アッセイが行われました。 AuNPのアポトーシス特性は、真核細胞のコメットアッセイでテストされ、最小限の副作用で治療に使用できる濃度を推定しました。したがって、本研究は、環境的および生物学的損傷を最小限に抑えるために、安全な範囲内で医療およびナノテクノロジーのインターフェースでAuNPの合成と適用を最適化することを試みています。
メソッド
菌類、バクテリア、植物の成長条件
Tricholoma crassum (バーク)サックナノ粒子の製造に使用されました。抗菌アッセイの場合、 E。コリ (DH5α)、アグロバクテリウムツメファシエンス (LBA4404)、 Eの多剤耐性(MDR)株。コリ (DH5α)、および A。 tumefaciens (LBA4404)を使用しました。植物病原菌 Magnaporthe oryzae および Alternaria solani に使われていた。トマト(品種プサルビー)とタバコ(品種SR1)の苗木は、16:8時間の明暗光周期、28±1°C、光強度50μmolm − 2 > s − 1 。
AuNPの合成
T。 crassum 菌糸体は、ポテトデキストロースブロス(PDB)で28°Cで7日間培養されました。 1gの菌糸体マットを10mlの脱イオン水と一緒にシェーカーで50RPM、28°Cで24時間、48時間、72時間攪拌しました。上澄みをワットマン濾紙No. 1.細胞ろ液(pH 5.2)を1 mMクロロ金酸水溶液(HAuCl 4 )とインキュベートしました。 )そして、菌糸体との24、48、および72時間のインキュベーションによって作成された細胞濾液の各タイプについて、レポート[2]に従って暗所で28°Cで1時間攪拌しました。
さまざまなpHでのAuNPの生合成では、1 MHClまたは1M NaOHを使用して、インキュベーション前に無細胞ろ液のpHを酸性範囲(3.5)およびアルカリ性範囲(7、8、および9)に調整しました。 AuNPは、さまざまな反応温度(0、15、28、75、および100°C)、さまざまな濃度の無細胞ろ液(×0.5、×1、×2)、およびクロロ金酸イオン(0.5、1、2 mM)を使用して合成されました。 。
UV-可視分光法
クロロ金酸溶液とともに1時間インキュベートした各細胞濾液からの上清の吸光度を、450〜750nmのUV-可視分光光度計を使用して分析し、吸光度スペクトルをプロットしました。
走査型電子顕微鏡(SEM)、透過型電子顕微鏡(TEM)、原子間力顕微鏡(AFM)、およびX線回折(XRD)
これらの分析は、Chowdhury etal。に従って行われました。 [15]いくつかの変更があります。 24時間の細胞濾液を使用して生成された金ナノ粒子と、それに続く1 mM HAuCl 4 との1時間のインキュベーション 28°C(pH 5.5)の溶液は、走査型電子顕微鏡(SEM)、透過型電子顕微鏡(TEM)、原子間力顕微鏡(AFM)、およびX線回折(XRD)を使用して特性評価されました。ガラススタブ上のAuNPの薄膜を真空乾燥し、FEI Quanta 200(FEI、USA)を使用してSEMにかけました。
AuNPの形状とサイズはTEMによって決定されました。 AuNP懸濁液の液滴(10μl)をカーボンコーティングされた銅グリッド上に置き、試料ホルダーにロードする前に真空乾燥にかけました。これらのナノ粒子のTEM顕微鏡写真は、低電圧(100 kV)構造のTECNAI GTEMを使用して取得されました。
AFMイメージングでは、AuNPを新たに劈開した白雲母Rubyマイカシート(Ruby Mica Co. Ltd.、インド)に堆積させ、真空乾燥機を使用して乾燥させました。音響交流(AAC)モードAFMは、Pico plus 5500 ILM AFM(Agilent Technologies、USA)を使用して実行されました。
XRD研究では、AuNP懸濁液の薄膜をスライドガラス上に均一に広げ、真空乾燥機を使用して乾燥させました。 XRDパターンは、電圧40 kV、電流40 mA、CuKα放射線(λ=1.54060/1.54443Å)で動作するD8 Advance DAVINCI XRDシステム(Bruker AXSPvt。Ltd。)で記録され、回折強度が記録されました。 35°から80°の2θ角度。
コンピューターソフトウェア分析
AuNPの測定とヒストグラムの作成は、OLYMPUSソフトウェアMEASUREITツールを使用して行われました。ナノ粒子の濃度は、Sriram etal。に従って計算されました。 [18]および私たちの以前の出版物、Chowdhury etal。 [15]。
多剤耐性を発生させるための細菌の形質転換
A。 tumefaciens LBA4404株および E。コリ DH5α株は、公開されているプロトコル[2、15]を使用して、プラスミドpCAMBIA2301およびpUC19とpZPY112をそれぞれ使用した形質転換により、多剤耐性になりました。
抗菌アッセイと細菌増殖アッセイ
これらのアッセイは、公開されているプロトコル[15]に従って行われました。 24時間の細胞濾液と1mM HAuCl 4 との1時間のインキュベーションを使用して合成された金ナノ粒子 28°C(pH 5.5)の溶液を、すべての生物学的アッセイに使用しました。ペーパーディスクアッセイでは、多分散AuNP(0.249、0.498、0.747、0.996、1.245μg)の量を増やして使用しました。各細菌株の新鮮な一晩培養物から、25μLのアリコートをLB寒天プレートに広げました。ナノ粒子溶液の希釈系列は、濃度31.121 mg / LのAuNP溶液と滅菌脱イオン水を使用して作成されました。 0.249、0.498、0.747、0.996、1.245μg(総量40μl)など、各ディスクの金ナノ粒子の量を増やした直径5 mmの滅菌紙ディスクを細菌プレートに置き、インキュベートしました。 A。 tumefaciens プレートを28°Cで48時間 Eでインキュベートしました。コリ 37°Cで12時間。 DH5αおよびLBA4404の増殖アッセイでは、7.5mlの細菌培養物に2.5mLのAuNPを添加しました。
抗真菌アッセイ
Mの水性懸濁液。 oryzae 胞子は6.1×10 5 で作られました 血球計算盤で胞子/ ml。この懸濁液の150μlをMEAプレートに広げました。多分散AuNP(0.249、0.498、0.747、0.996、および1.245μg)の量を増やした直径5 mmの滅菌紙ディスクをプレートに置き、28°Cでインキュベートしました。抑制ゾーンは2日後に測定されました。
AuNPで処理された細菌細胞の抗菌アッセイ
一晩液体LBA4404培養物を、等量のAuNP懸濁液(15.56 mg / L)で28°Cで12時間処理しました。 50μlの懸濁液を等量の0.4%トリパンブルー溶液(0.5 gトリパンブルー、500 mlグリセロール、450ml蒸留H 2 )と混合しました。 O、50 ml HCL)を複合顕微鏡(Leica DMLS、ドイツ)で観察しました。
AuNPの存在下での真菌胞子発芽アッセイ
ナノ粒子の希釈系列は、20、40、60、80、および100%の v で作成されました。 / v 濃度15.56mg / Lのナノ粒子のストック溶液を使用して、水で最終容量を100μlにします。これらの懸濁液の等量を50μlの Alternaria solani に加えました。 胞子懸濁液(4.2×10 5 胞子/ ml)、28°Cで培養。胞子は、複合顕微鏡(Leica DMLS、ドイツ)で0、2、4、6時間後に観察されました。
コメットアッセイ
AuNPのアポトーシス誘発特性は、いくつかの変更を加えた標準的なコメットアッセイ[19]によって測定されました。タバコまたはトマトの葉は、増加する濃度(0、15、20、および30% v )にさらされました。 / v タバコ用; 5、10、15、および20% v / v トマトの場合)24時間のAuNP(15.56 mg / Lストック)。孤立した核の電気泳動は、0.74 V / cm(25 V、300 mA)で4°Cで30分間実施しました。スライドを0.4M Trisバッファーで中和し、メタノールで脱水し、臭化エチジウム(20μg/ ml)で染色し、515〜560nmの励起フィルターと590nmのバリアフィルターを備えた蛍光顕微鏡で観察しました。データはTritekCometScoreソフトウェアで分析されました。
SDS-PAGE
私たちの出版物[15]に従ってタンパク質を単離しました。ナノ粒子に結合したタンパク質を分析するために、AuNPを滅菌水で洗浄し、Laemmliバッファーで10分間煮沸し、8000rpmで10分間遠心分離しました。 SDS処理および未処理のサンプルを12%SDS-PAGEで泳動しました。
がん細胞株の培養
肉腫180細胞株は、10%ウシ胎児血清、200 U / mlペニシリン、および200μg/ mlストレプトマイシンを含むRPMI1640培地[20]で、37°C、5%CO 2 で培養されました。 加湿インキュベーター内。
プラスミドDNA-AuNP複合体の癌細胞への送達
公開されているプロトコル[15]を使用して、pCAMBIA1302を含むDH5αからプラスミドを単離しました。 gfp のコンストラクトを含むプラスミド pCaMV35sプロモーター下でクローン化された配列は、標準プロトコルを使用して肉腫180細胞に送達されました[21]。 GFPに特異的なフィルター(励起最大=395 nm)(Axioskop-40、Carl Zeiss)を使用して、蛍光顕微鏡下で細胞を観察しました。裸のプラスミドDNAで処理された細胞はコントロールとして保持されました。
溶血性アッセイ
溶血アッセイは、標準的なプロトコル[22]に従って行われました。等量の赤血球(1.6×10 9 赤血球/ mL)をさまざまな濃度のAuNP(0.1、0.5、1、5、10、および20μl/ mL(ストック15.56 mg / Lから))で1時間処理し、さまざまな濃度でのナノ粒子の溶血活性を計算しました。
結果と考察
pH5.5でのAuNPの生合成
合成中、AuNPの形成は、表面プラズモン共鳴(SPR)の変化による細胞濾過液の色の淡黄色から紫色への変化によって示されます[23]。 1 mM HAuCl 4 真菌のろ液を含まない溶液は緑がかった黄色であり(図1a、「A」)、 Tの細胞ろ液でした。 crassum これは淡黄色でした(図1a、「B」)。インキュベーション後、混合物の色は1時間以内に紫色に変化し(図1a、「C」)、AuNPの形成を示しています。
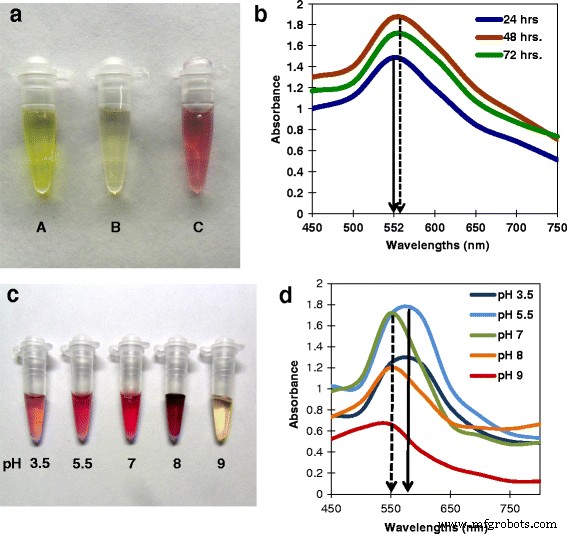
Tricholoma crassum を使用したAuNPの生合成 および分光分析。 a 反応中の色の変化。 A HAuCl 4 の20mM溶液 。 B Tricholoma crassum の菌糸体を含まない細胞ろ液 。 C 1 mM HAuCl 4 24時間の細胞ろ液を1時間使用すると、AuNPの合成を示す紫色が表示されます。 b さまざまなインキュベーション期間(24、48、72時間)で合成されたAuNPのUV-visスペクトル。実線の矢印は、24時間のインキュベーション期間における552nmでの吸収ピークを示しています。破線の矢印は、インキュベーション期間の増加に伴う吸収極大のわずかな赤方偏移を示しています。 c 異なるpHで合成されたAuNPは、異なる色を示します。 d 同じのUV-visスペクトル。実線の矢印は、pH5.5の552nmでの吸収ピークを示し、破線の矢印は、pHの増加に伴う吸収極大の青方偏移を示します
微生物からナノ粒子を生成する他の方法の1つの欠点は、これらが時間がかかり、生物が毒素を生成することです。以前は、 Phanerochate chrysosporium 無細胞抽出物を利用して、90分でAuNPを生成しました[3]。最近、銀と金のナノ粒子が Sporosarcinakoreensis を使用して生合成されました 1〜2日の延長された反応時間を使用します[24]。レポートでは、AuNPの生産時間が1時間に短縮されました。
紫外から可視分光法
金属ナノ粒子の光吸収スペクトルは、形状、凝集、およびSPRによって支配され、これらは粒子のサイズと形状に応じてシフトします[25、26]。図1bは、24、48、72時間にわたって生成された細胞濾液とそれに続く1 mM HAuCl 4 との1時間のインキュベーションを使用して合成されたAuNPのUV-visスペクトルを示しています。 溶液(pH 5.5)。ここでは、24時間の細胞ろ液の吸光度のピークは552nmにあります。 552 nmで最初に現れる横方向プラズモン共鳴バンドは、24時間から48時間および72時間にわずかに赤方偏移し(図1bの実線と破線で示されています)、吸光度の強度が徐々に増幅された赤方偏移を確認します。ピークの広がりは、粒子が多分散していることを示しています。約を中心とした強力なSPR。 550〜560 nmは、金コロイドの典型です(図1b)。潜伏期間(24、48、72時間)の増加に伴って細胞ろ液の濃度が増加すると、それに比例して吸光度も増加しました。反応を28°Cで行った場合、1時間後に平衡に達し、30日間安定しており、おそらく安定化タンパク質コートが原因で凝集の形跡はありませんでした。以前の研究では、BSAでコーティングされたAuNPは凝集を示さなかった[27]。さらなる研究は、24時間の細胞ろ液で調製されたAuNPを使用して行われました。
さまざまなpHおよびUV-vis分光法でのAuNPの生合成
AuNPは、3.5、5.5、7、8、および9のさまざまなpHで合成され、ピンクから濃い紫色になりました(図1c)。 UV-vis分光法は吸光度の最大値を示し、SPR波長はpH3.5からpH5.5に増加し、赤方偏移をもたらしました。ただし、pHを7からpH 9にさらに上げると、最大吸光度とSPR波長が減少し、青方偏移を示します(図1d)。 pH 9のピークは振幅が小さく、色も変化していません。これは、少量のAuNPのみが形成されたことを示しています。これは酵素生合成であるため、pH 9はおそらくAuNPの形成に必要な酵素反応を阻害しました(図1d)。異なるpHで生成されたナノ粒子は、室温で1か月後に凝集しませんでした。
さまざまな温度、基質濃度、および前駆体濃度を使用したAuNPの生成
生合成中の物理化学的条件の最適化は、機能的に効率的なナノ粒子の生成にとって重要です[28]。より高濃度のろ液を使用した合成では、より多くの色強度と最大吸光度を持つAuNPの生成が増加しました(図2a、b)。最大局在表面プラズモン共鳴(LSPR)のわずかな青方偏移が、×2ろ液での合成で観察され、粒子間距離の増加とクラスターサイズの減少を示しています[29、30]。
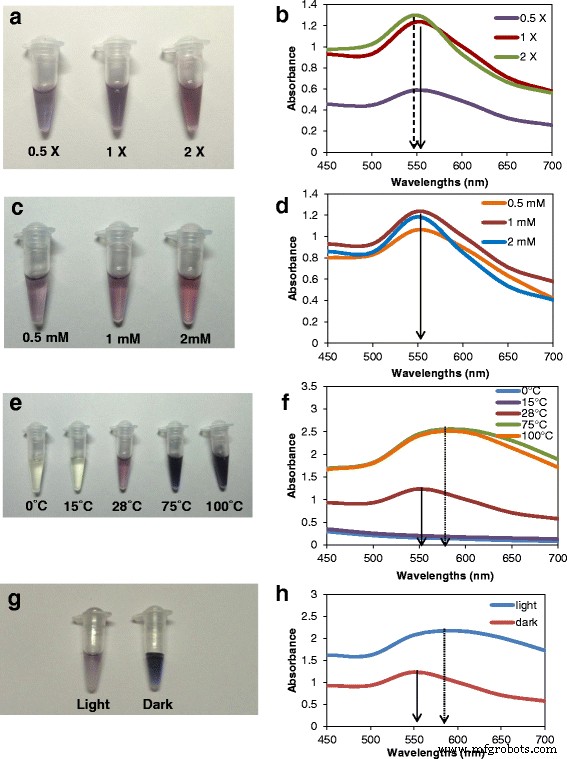
Tricholoma crassum からのAuNPの生合成とUV-vis分光法 異なる合成パラメータを使用します。 a 、 b 異なる濃度の細胞ろ液。 c 、 d さまざまな濃度のHAuCl 4 。 e 、 f 異なる反応温度。 g 、 h 暗闇と光の下で。実線の矢印は、×1の細胞ろ液と1 mM HAuCl 4 の552nmでの典型的な吸収ピークを示しています。 28°CのpH5.5で、破線の矢印は吸収極大の青方偏移を示し、点線の矢印はSPRバンドの赤方偏移を示します
テストした塩化金の3つの濃度(0.5、1、および2 mM)のうち、合成は1 mMで最大であり、吸収は552nmで最大でした。 (図2c、d)。合成には28°Cが最適であることがわかりました。より高い温度(75および100°C)は、AuNPのより速い合成を仲介しましたが、暗い色と最大吸収の赤方偏移(図2e、f)は、28°Cよりも大きな粒子を示しました。 0および15°Cでは、550〜600nm内に発色または吸光度のピークはありませんでした。光で合成すると、濃い紫色になり(図2g)、吸収極大の赤方偏移が大きくなり、ピークが広くなり(図2h)、粒子が大きくなります。粒子のサイズと数はDLSで分析されました(図3a–j)。ナノ粒子のサイズと凝集のこの変化は、関連する有機物の性質と量に依存し[30]、これも合成条件によって異なります。
合成されたAuNPのサイズ分布を示すDLS a ×1無細胞ろ液、1 mM HAuCl4、28°C、暗所、 b 光の下で、 c ×0.5無細胞ろ液、 d ×2無細胞ろ液、 e 0°Cで、 f 15°Cで g 75°Cで h 100°Cで i 0.5 mM HAuCl 4 および j 2 mM HAuCl 4
走査型電子顕微鏡(SEM)および透過型電子顕微鏡(TEM)
SEMは、倍率80000倍で、サイズが小さく、幾何学的形状が異なるAuNPを示しました(図4a)。 TEM分析では、直径2〜22 nmのサイズ範囲の多分散AuNPが明確に示されました(図4b)。サイズ分布グラフは、サイズ範囲5〜10 nmのAuNPが最も高い周波数であり、2〜5 nm、10〜15 nm、15〜20 nm、および20〜22 nmが続いていることを示しています(図4c)。これらは、直径5 nm以下の小さな円形または菱形、六角形、立方体、二等辺三角形、辺が4.36〜22.94 nmのほぼ正三角形など、さまざまな幾何学的形状でした(図4d、e)。
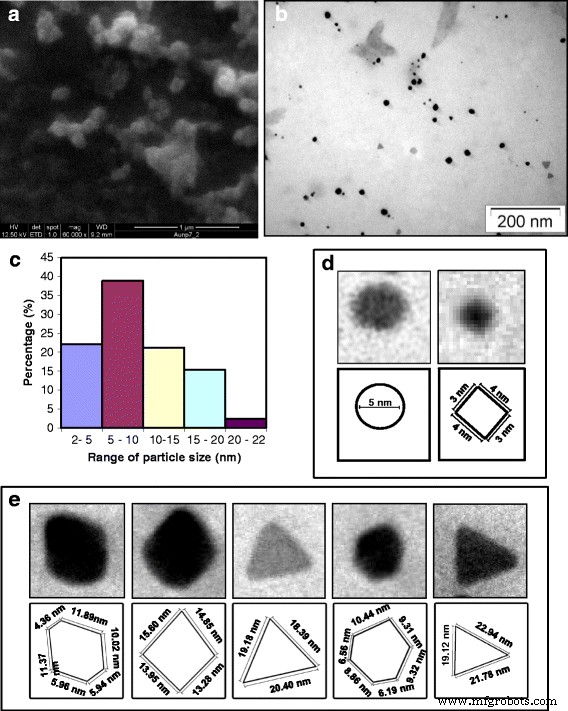
AuNPの電子顕微鏡。 a 倍率×80,000でのSEM。 b 顕微鏡視野で異なるサイズの分散粒子を示すTEM。 c AuNPの粒度分布を示すヒストグラム。個々のAuNPの拡大図で、さまざまな幾何学的形状とそれらの寸法による図式表現を示しています。 d サイズ範囲が小さい球形(左)と菱形(右)。 e 左から:六角形、正三角形、菱形、多面体、二等辺三角形
原子間力顕微鏡(AFM)
図5aは、分散したAuNPのAFM画像を示しています。 2次元ビュー(図5a)は、粒子の表面の厚さがほぼ同じであることを示しています。これらの粒子の高さは、単一表面の3D AFMで視覚化されました(図5b)。ランダムな線形ゾーン(点線でマーク、図5c)上にあるAuNPのAFM 2Dグラフは、1〜4nmの範囲の高さを示しています。平面プラズモンバンドは、粒子の厚さがエッジの長さよりも小さいことを示していました。
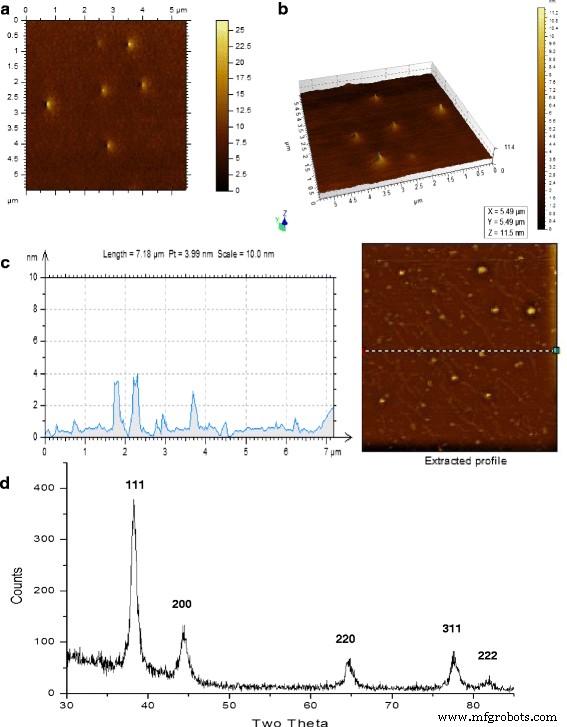
AuNPのAFMおよびX線回折。 a AFM画像:上面図。 b AFM画像:3次元ビュー。 c AuNPのAFM画像とフィールドの点線上にあるナノ粒子の高さのグラフプロファイル。 d 金に典型的なピークを示すナノ粒子のX線回折パターン
X線回折研究(XRD)
粒子の薄膜のXRDパターンは、AuNPのみの存在を明らかにしました。約38°の強い回折ピークは、一般に、面心立方(FCC)構造の{111}ファセットに起因します[3]。ここでのXRDパターンは、FCC構造に起因する38.23に支配的な回折ピークを示しました。他の4つのファセットの回折ピークは弱かった。 38.23、44.31、64.60、77.58、および81.63の4つの異なるブラッグ回折ピークは、AuNPのピークと厳密に一致していました(図5d、追加ファイル1:表S1)。
AuNPの濃度の計算
ナノ粒子の濃度[15]は、1 mM HAuCl 4 とインキュベートした24時間の細胞ろ液で生成された粒子で15.56mg / Lであることがわかりました。 。
病原菌と真菌を使用したAuNPの抗菌アッセイ
AuNPsは、人間の細菌だけでなく、植物の病原性細菌や真菌に対して強力な抗菌活性を示しました。ナノ粒子の抗菌活性は、AuNPの量を増やして、つまり0.249、0.498、0.747、0.996、および1.245μgの紙ディスクを使用して分析しました。ヒトバクテリア E。コリ (DH5α)、植物病原菌 A。 tumefaciens (LBA4404)および植物病原菌 M。 oryzae に使われていた。 AuNPは、最低濃度でもこれらすべての微生物を阻害し、阻害ゾーンは粒子濃度の増加に比例して増加しました(図6)。図6a–cは、 Eの抑制ゾーンを示しています。コリ (DH5α)、 A。 tumefaciens (LBA4404)および M。 oryzae 、 それぞれ。図6f–hは、使用されたAuNPの量の関数としてのこれら3つの微生物の阻害ゾーンのグラフを示しています。真菌抽出物だけでは阻害効果はありませんでした。 3つの微生物の阻害の比較傾向は、LBA4404および<i> Mの阻害効果と比較して、DH5αに対するより大きな阻害効果を示している。 oryzae (追加ファイル2:図S1a)。
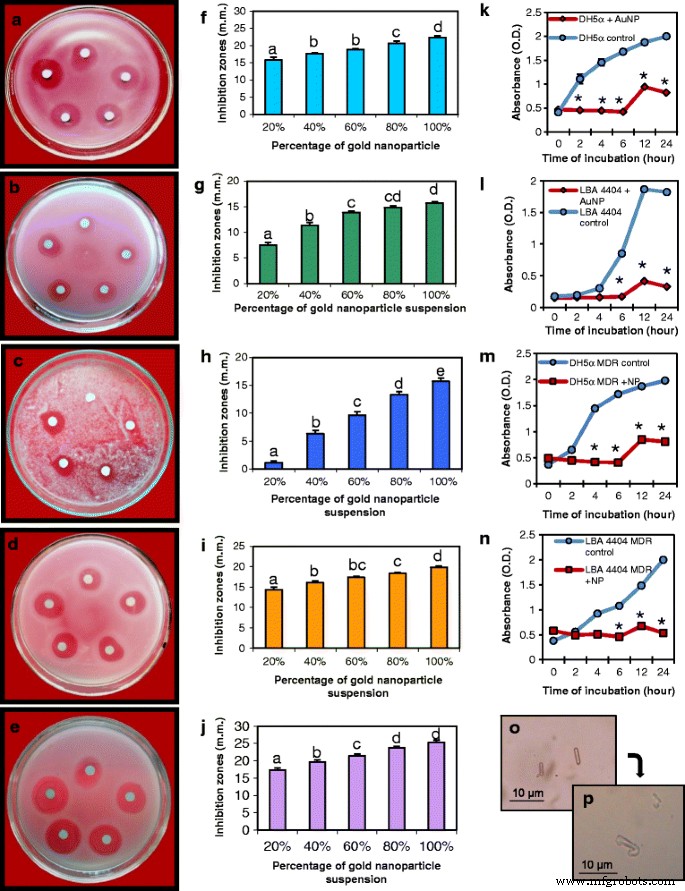
病原性細菌、真菌、および多剤耐性(MDR)細菌に対するAuNPの抗菌特性のアッセイ。 a 、 f Eの阻害ゾーンが増加するナノ粒子のディスク拡散アッセイを示すプレートおよび対応するグラフ。コリ 。 b を使用した同様のアッセイで得られた阻害ゾーン 、 g アグロバクテリウムツメファシエンス 。 c 、 h イネいもち菌 。 d 、 i MDR E。コリ 。 e 、 j MDR A。 tumefaciens 。すべての実験は、紙のディスク上のAuNPの量を増やして行われました。上から時計回りに:0.249μg(20%)、0.498μg(40%)、0.747μg(60%)、0.996μg(80%)、および1.245μg(100%)のAuNP。データは、3回の繰り返しの平均±SEです。異なる文字は、サンプル間の統計的に有意な差を示します( P <0.05、テューキーのHSD検定)。 k の成長曲線に対するAuNPの影響 E。コリ 、 l A。 tumefaciens 、 m MDR E。コリ 、および n MDR A。 tumefaciens 。アスタリスクは、制御するための重要な違いを示します(学生の t テスト、 P <0.05)。 o コントロールの顕微鏡検査 A。 tumefaciens 細胞。 p A。 tumefaciens AuNPsによる治療後の細胞の完全性の喪失を示す
多剤耐性のヒトおよび植物の病原菌を使用したAuNPの抗菌活性の分析
耐性遺伝子を持つプラスミドを運ぶ多剤耐性(MDR)DH5αとLBA4404が作られました。 pUC19を運ぶMDRDH5αは、100μg/ mlのアンピシリンと35μg/ mlのクロラムフェニコールに耐性がありました。 pCAMBIA2301で形質転換されたLBA4404は、25μg/ mlのリファンピシンと50μg/ mlのカナマイシンに耐性がありました。 AuNPは、MDRDH5αおよびMDR LBA4404に対して強力な阻害活性を示しました(図6d、e)。図6i、jは、ナノ粒子の濃度の増加に伴う阻害の増加を示すグラフです。 2つのMDR細菌の阻害の比較傾向は、 Aの阻害ゾーンが大きいことを示しています。 tumefaciens E.coli のそれと比較して (追加ファイル2:図S1b)。
AuNPの存在下での経時的な細菌増殖アッセイ
AuNPで処理されたDH5αの増殖曲線は、コントロールセットとは大幅に異なっていました(図6k、m)。 DH5αとMDRDH5αのコントロールセットでは、増殖曲線の対数期は接種から2時間以内に始まりましたが、AuNPで処理したDH5αとMDRDH5αでは、接種後6時間まで増殖は観察されませんでした。増殖曲線は、対照細胞と処理細胞の両方で12時間後に定常期に達しましたが、処理細菌の場合、増殖は大幅に減少しました。 LBA4404の増殖曲線は、AuNP処理セットでは6時間であったのに対し、コントロールセットでは接種後4時間で対数増殖期の開始を示しました。 MDR LBA4404の成長曲線に対する同様の効果が観察されましたが、コントロールMDR LBA4404では、対数期が2時間以内に開始しました(図6l、n)。
細菌細胞の形態と生存率に対するAuNPの影響
一般に、ほとんどのナノ粒子は細胞膜に効率的に付着し、吸着され、その後細胞の完全性に影響を与える可能性があります[31]。通常の A。 tumefaciens バクテリアは輪郭がはっきりした棒状です(図6o)。 AuNPとインキュベートしたときの細菌は、形態が歪んでおり、外膜が崩壊して輪郭が不規則になり、細胞の形状とサイズの完全性が失われていました(図6p)。これは、以前に Eで観察されたものと一致しています。コリ シリカナノ粒子で処理された細胞[32]。バクテリア細胞をナノ粒子と長期間インキュベートすると、細胞が完全に崩壊することがわかりました。
真菌胞子発芽アッセイ
AuNPは、植物病原菌 Alternaria solani の病原性の強力な抑制因子でした。 。さまざまな潜伏期間でAuNPの投与量を増やして真菌分生子を処理すると、発芽頻度と発芽管の長さが徐々に減少することが示されました(図7a)。分生子の代表的な写真(図7a)とグラフは、菌類の分生子から出てくる発芽の割合(図7b)と発芽管の平均の長さ(図7c)が、粒子の投与量の増加とインキュベーションの増加とともに減少したことを示しています期間。したがって、これらのAuNPは、胞子の発芽の抑制と菌糸の成長の遅延によって媒介される重要な抗真菌特性を持っていました。
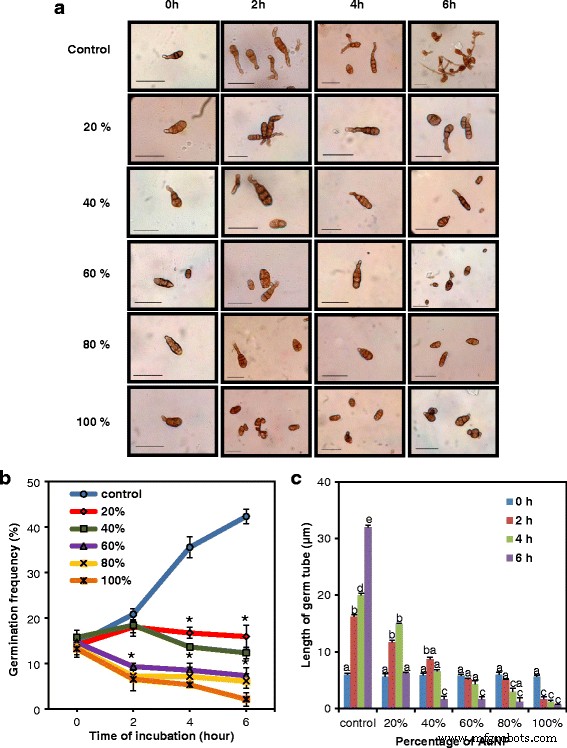
植物病原菌 Alternaria solani の胞子発芽に対するAuNPの影響; a AuNPで処理した後の胞子(bar =30μm)。上から下の行:コントロール胞子に続いて、さまざまな希釈のAuNP(15.56 mg / Lストック)で処理された胞子。左から右の列:AuNPとのインキュベーション時間が長くなり、100%AuNPとの6時間のインキュベーションで発芽が最小になります。 b インキュベーション時間の関数としてのAuNPの異なる濃度での胞子発芽頻度(%)。アスタリスクは、制御するための重要な違いを示します(学生の t テスト、 P <0.05)。 c インキュベーション時間の関数としてのAuNPの異なる濃度での平均生殖管長。データは、3回の繰り返しの平均±SEを表します。異なる文字は、サンプル間の統計的に有意な差を示します( P <0.05、ダンカンの多重範囲検定)。胞子の発芽頻度と平均発芽管長は、AuNPの投与量の増加と潜伏期間の増加とともに減少しました
したがって、これらのAuNPは、細菌や真菌に対して抗菌機能を持っていますが、AuNPではほとんど報告されていません。多剤耐性のヒト細菌に対して毒性のあるこれらのAuNPは、MDRまたは治療が困難な広範囲の薬剤耐性(XDR)細菌関連のヒト疾患の治療に利用できます。さらに、アグロバクテリウムのような典型的な植物病原体に対する抗菌効果 そして菌類、それは粒子をナノ農薬の形で殺菌剤および殺菌剤として適したものにします。これらは、土壌の健康の悪化、農業生態系の劣化、環境汚染、および病原体の耐性を引き起こす農薬の過剰かつ無差別な使用を排除することにより、作物損失の持続可能な管理を可能にします[33]。
AuNPには強力なアポトーシス誘発特性があります
単一細胞ゲル電気泳動アッセイまたはコメットアッセイは、材料のアポトーシス効果を比較するための高感度な方法です[34、35]。タバコの葉の細胞を7.78mg / LのAuNPで15、20、30分間処理すると、アポトーシスが徐々に増加し、尾のDNAの割合が増加しました(図8a、「A」、「B」)。 、 'CD')。 DNAの最大移動は、処理の30分後に発生し、テールDNA値は28.44±0.74%であり、未処理のコントロール細胞よりも大幅に高かった(1.7±0.59%)。 15分と20分のインキュベーション期間ではDNAの移動が少なくなり、テールDNA値はそれぞれ7.25±2.56と19.19±1.54%でした(図8b)。したがって、これらのAuNPは、高用量で真核細胞にアポトーシスを誘導する能力を持っています。最近、ナノ粒子は、癌細胞を標的にして殺すための新しい戦略で使用されています。動的かつ定量的なイメージングで示されているように、癌の代替療法としてのナノ粒子の適用の成功は、粒子のアポトーシス特性に依存します[36]。したがって、現在の発見は、これらのAuNPの強力なアポトーシス誘発特性を示しており、将来の癌治療に有望です。
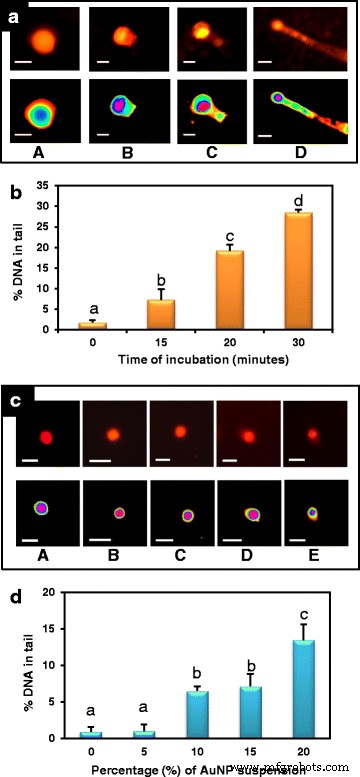
AuNPのアポトーシス特性のアッセイ。 a A のAuNPで処理されたタバコの葉の核のコメットアッセイと蛍光顕微鏡画像 0分 B 15分、 C 20分、 D 30分(バー=5μm)。コントロールセット(0分間のインキュベーション)では、ほとんどのDNAが彗星の頭にありますが、より長い処理を受けた細胞は、DNA損傷の増加と、より長い彗星の尾を示します。 b 異なるインキュベーション期間後の%テールDNA±SEの平均。データは、3回の繰り返しの平均±SEを表します。異なる文字は、サンプル間の統計的に有意な差を示します( P <0.05、テューキーのHSD検定)。 c A で処理されたトマト葉細胞の核の画像を示すアポトーシスに対するAuNPの閾値投与量のアッセイ 0%(コントロール)、 B 5%、 C 10%、 D 15%、 E 24時間のAuNP懸濁液の20%(bar =10μm)。 d 異なる濃度のAuNPで24時間処理した後の平均%テールDNA±SE。データは、3回の繰り返しの平均±SEを表します。異なる文字は、サンプル間の統計的に有意な差を示します( P <0.05、テューキーのHSD検定)。 10%未満の投与量はごくわずかなDNA損傷を示し、20%を超える投与量はDNA損傷の開始を示します
これらのAuNPは低用量では非アポトーシス性です
治療薬としての金属ナノ粒子の安全な適用には、境界毒性における粒子の生物学的効果の事前決定が必要です[37]。アポトーシスに必要なナノ粒子のしきい値レベルは、トマトの葉の細胞をさまざまな濃度のAuNPで24時間処理することで得られました(図8c)。低濃度、たとえば5% v / v AuNPストック懸濁液(15.56 mg / L)は、24時間の曝露後でも、トマト細胞に対して有意なアポトーシス誘発効果を示しませんでした。 10%AuNP処理後の核は、未処理の対照核(尾部のDNAの0.95±0.66パーセント)および5%AuNP懸濁液で処理された核(尾部のDNAの1.04±0.89パーセント)よりも高い6.52±0.63パーセントの尾部のDNAを示しました)。トマトの葉の細胞を15%および20%のAuNPで処理すると、DNA損傷がわずかに上昇し、尾部のDNAの7.15±1.70および13.47±2.16パーセントを示し、有意なアポトーシス誘発効果を示しました(図8d)。したがって、低用量では、アポトーシス誘発効果は無視できる程度であり、これらのナノ粒子が真核細胞で抗菌剤または薬物/遺伝子送達媒体として使用される可能性があります。
AuNPはタンパク質でコーティングされています
以前のいくつかの研究では、天然由来のタンパク質でコーティングされたナノ粒子について言及されています[15]。低倍率のTEMは、AuNPがタンパク質のような物質に囲まれていることを示しました(図9a、b)。材料の性質を確認するために、AuNPを洗浄し、他のレーンの無細胞抽出物と一緒にSDS-PAGEで泳動しました(図9c)。 SDSでの煮沸は、表面に結合したタンパク質をナノ粒子から分離するのに役立ちました。煮沸したナノ粒子(レーン4)は、レーン2(細胞ろ液)に存在するタンパク質バンドと同様の40kDaの単一の強いバンドの存在を示しました。ただし、煮沸されていないサンプル(レーン3)では、AuNPに結合したかすかなタンパク質バンドが116kDaレベルで見られました。キャッピングタンパク質が粒子から解離したため、40kDaレベルで別のかすかなバンドが現れました。タンパク質コートは、溶液中のナノ粒子の安定性とその触媒活性を促進することが知られています[28、38]。ナノ粒子の周りに自然に形成されたタンパク質コートは、DNAまたは疎水性薬物の容易な吸着と送達を含む生物医学的使用のためにそれらを機能的に効率的にします[39]。ペプチドおよびAuNPのタンパク質支援送達は、中枢神経系障害の治療における血液脳関門を克服するために首尾よく使用されてきました[14]。したがって、これらの生体適合性AuNPは、サイズが小さく、独自の物理化学的特性やその他の利点があるため、いくつかの生物医学的用途に適している可能性があります。
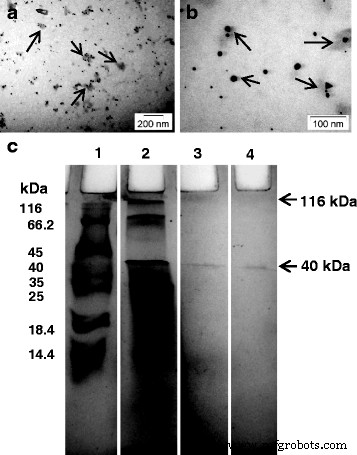
プロテインキャップ分析。 a 、 b AuNP周辺のキャッピングタンパク質層(矢印)を示すTEM画像。 c Tから分泌された細胞外タンパク質のSDS-PAGE。 crassum ナノ粒子に関連するタンパク質。レーン1、分子サイズマーカー。レーン2、総細胞外タンパク質。レーン3、沸騰せずにロードされたナノ粒子は、116kDaマークでAuNPに結合したかすかなタンパク質バンドを示しています。 40kDaマークに分離したタンパク質のバンドもあります。レーン4、1%SDSローディングバッファーで煮沸した後のナノ粒子は、116kDaバンドと明確な40kDaバンドの消失を示しています。矢印は40kDaを示します
AuNPは、緑色蛍光タンパク質(GFP)遺伝子を肉腫180がん細胞株に送達する可能性があります
gfp を含むプラスミドDNApCAMBIA1302 AuNPsと複合体を形成したマーカー遺伝子を使用して肉腫180細胞を治療しました。細胞は緑色の蛍光を発し、プラスミドDNA / AuNP複合体の取り込みとそれに続く癌細胞での遺伝子の発現を示しましたが、裸のプラスミドDNAのみで処理した細胞は蛍光を示しませんでした(図10a、b)。これは、これらの粒子が癌細胞に遺伝子を送達するだけでなく、遺伝子が安定して発現され、細胞に送達されると機能を維持する可能性が高いことを裏付けています。
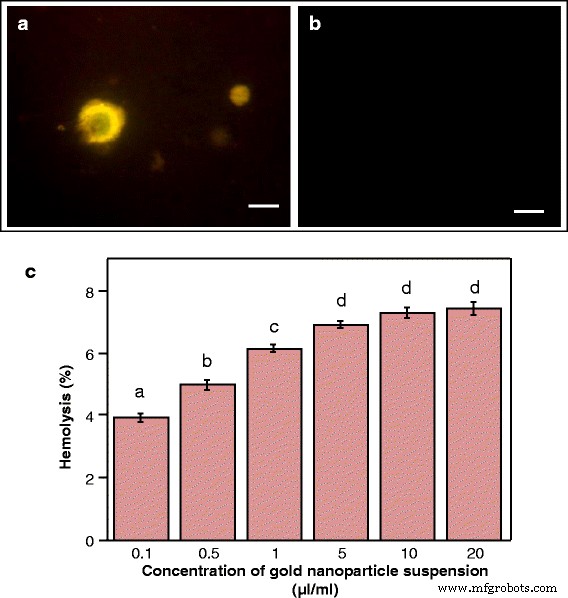
肉腫180癌細胞へのAuNPを使用した遺伝子送達およびヒト赤血球を用いた溶血アッセイ。 a の蛍光顕微鏡画像 DNA-AuNP複合体および b の取り込み後に緑色蛍光タンパク質を発現する癌細胞 遊離プラスミドDNAで処理されたコントロール細胞(バー=20μm)。 c AuNPのさまざまな希釈による溶血の割合。データは、3回の繰り返しの平均±SEを表します。異なる文字は、サンプル間の統計的に有意な差を示します( P <0.05、テューキーのHSD検定)
初期の金属ナノ粒子は、網膜血管新生、HIV、ダルトンリンパ腫などのさまざまな疾患の治療において計り知れない治療の可能性を示し、肝炎ウイルス、呼吸器合胞体ウイルス、単純ヘルペスウイルスに対して活性を示しました[18]。これらの報告と一致して、AuNPを使用した現在の研究における金ナノキャリアベースの薬物送達は、診断、薬物送達、および癌治療を含む多くの医療用途における有望なメディエーターと見なすことができます。
AuNPとヒト赤血球および毒性アッセイとの互換性
赤血球は細胞膜システムのシンプルで便利なモデルであり、ナノ粒子と膜の相互作用の研究に使用されます[40]。溶血アッセイは、さまざまな濃度でのAuNPの膜溶解活性を解明します。図10cは、ナノ粒子の膜溶解活性が低濃度で無視できることを示しています。高濃度のナノ粒子懸濁液(最大20μl/ mL)で見られる最高の溶血活性は8%未満であり、これは血液毒性が非常に低いことを示しています。 AuNPの濃度の増加に伴って溶血活性が徐々に増加するのは、赤血球とのより多くの粒子の親和性と接着の増加による可能性があります。粒子の細胞膜へのこの親和性は、それらの細胞輸送を促進すると予想される。溶血活性が低く、細胞への取り込みが効果的であるため、ナノ粒子は安全で効率的な治療薬の開発に非常に適しています[41]。
結論
食用菌根菌を使用して、天然のタンパク質コートとの短い反応時間でさまざまな幾何学的形状のAuNPを合成することで、この方法はシンプルでユニークなものになります。食用キノコに由来するタンパク質コートは、感知できるほどの毒性作用はなく、粒子の表面へのDNAの容易な付着に有利に働きます。全体として、これらのAuNPは、癌細胞への遺伝子送達のための抗菌性のアポトーシス剤として有望です。
糸状菌は流れの圧力や攪拌に耐えることができるため[15]、 T。 crassum 発酵槽で培養して、無毒の農業廃棄物を使用して大規模にAuNPを生成できるため、製品やシステムの補充オプションを簡単に回収できます[2]。 AuNPはさまざまな幾何学的形状であるため、遠心分離などの機械的手段を備えたスコープ形状ベースの品揃えがあり[42]、特定の形状ベースの特性に応じて利用できます。
ナノマテリアル
- 化学療法センサー用の金ナノ粒子
- 改善された診断および治療用途のための多機能金ナノ粒子:レビュー
- 癌治療のためのナノ粒子:現在の進歩と課題
- コバルトをドープしたFeMn2O4スピネルナノ粒子の調製と磁気特性
- ICAをロードしたmPEG-ICAナノ粒子の調製とLPS誘発性H9c2細胞損傷の治療におけるそれらの応用
- HT29およびSPEV細胞株に対するAuナノ粒子の影響のinvitro研究
- フェニルトリメトキシシランで修飾されたアルミナナノ粒子をベースにしたAl2O3:SiOCナノコンポジットの形成と発光特性
- 6-メルカプトプリンとニューロン透過性ペプチドで修飾された金ナノ粒子によるSH-SY5Y細胞増殖の促進
- 金および銀ナノ粒子のグリーン合成のためのPlatycodiRadix(Platycodon grandiflorum)からのPlatycodonサポニン
- ゾルゲル法で合成されたPrドープペロブスカイトマンガン酸塩La0.67Ca0.33MnO3ナノ粒子の微細構造、磁気、および光学特性
- ポリマーナノコンポジットの界面/相間特性および引張強度に及ぼすナノ粒子のサイズおよび凝集/凝集の影響