


蛍光ネオ糖タンパク質金ナノクラスター:合成と植物レクチンセンシングおよび細胞イメージングへの応用
要約
炭水化物とタンパク質の相互作用は、受精、細胞シグナル伝達、宿主と病原体のコミュニケーションなどの基本的な生物学的プロセスを仲介します。ただし、グリカン認識イベントは非常に複雑であるため、近年、それらの分析またはアプリケーションを可能にする新しいツールが登場しています。ここでは、標的分子として二分岐N型糖鎖G0、担体/モデル抗原として卵白アルブミン、イメージングプローブ(G0-OVA-AuNCs)として蛍光金コアを含む、ネオ糖タンパク質で機能化された蛍光金ナノクラスターの最初の調製について説明します。続いて、生成されたG0-OVA-AuNCが、植物レクチンの特異的センシングおよび樹状細胞のinvitroイメージングに有用であることを示します。
はじめに
10から100個の金の原子によって形成される金ナノクラスター(AuNC)は、その魅力的な化学的および物理的特性により、科学界の注目を集めています[1]。 3 nm未満の金ナノクラスターは、電子のフェルミ波長に近づき、サイズに依存する蛍光発光を引き起こし、invitroおよびinvivoアプリケーションのセンシングおよびイメージングプローブとしての機会を提供します[2、3、4]。現在の蛍光アッセイは、主にローダミンやフルオレセインなどの有機色素、またはあまり一般的ではない量子ドットを使用しています[5、6、7]。ただし、一部の有機色素の光化学的安定性が低い、pHに敏感な蛍光、または水溶性が低い、量子ドットの毒性があるため、それらの使用は危険にさらされる可能性があります[8]。この文脈では、金ナノクラスターは、上記の制限がない代替の超小型フルオロフォアと見なすことができます。さらに、AuNCは、大きなストークスシフトと、可視赤色から近赤外(IR)領域への蛍光発光波長を特徴としています。これは、組織の透明度ウィンドウと重なるため、バイオイメージングに非常に適しています[8,9,10]。
金ナノクラスターのタンパク質支援合成は、ウシ血清アルブミン(BSA)を使用して2009年に最初に報告され[11]、それ以来、水溶性タンパク質保護ナノクラスターはナノサイエンスの新たなトレンドになりました[9、12、13]。 AuNCの調製のための糖タンパク質[14]にはあまり注意が払われておらず、合成ネオ糖タンパク質からの足場としてのAuNCの形成を説明している報告はありません。一般に、グリコシル化は、糖タンパク質の物理化学的特性、例えば、折り畳み、循環寿命、または安定性を調節します。また、受容体-リガンド認識など、タンパク質の重要な生物学的機能にも影響を及ぼします。したがって、タンパク質からの合成ネオ糖タンパク質の生成は、注意深く設計および特性評価された炭水化物の化学的付着によって[15]、生物学的用途のための新しい機能特性を備えさせることができます。炭水化物は炭水化物結合タンパク質との相互作用を通じて多数の異なる生物学的プロセスに関与するため[16、17]、invitroおよびinvivoの両方で炭水化物認識イベントを研究および活用するための新規プローブとしてネオ糖タンパク質金ナノクラスターを想定しています[18]。この点で、タンパク質表面に複数のグリカンコピーが存在すると、炭水化物の多価提示が提供され、その後、結合親和性が向上します。
ここでは、植物レクチンのセンシングプローブとして、および樹状細胞(DC)のin vitroイメージングのためのレクチン受容体のターゲティング試薬として、自己蛍光ネオ糖タンパク質で機能化されたAuNCの使用を検討します。レクチンは、さまざまな生物学的認識現象を支援する炭水化物結合タンパク質です。たとえば、高等植物では、レクチンは外来糖タンパク質の認識と凝集によって植物を食べる生物を防ぎ、それらの成長と増殖を阻害します[19]。一方、哺乳類のDCでは、C型レクチン受容体(CLR)が細胞表面に発現し、病原体の認識に大きな役割を果たします[20]。グリカンで装飾された抗原は、特定のCLRによって認識され、さらにエンドサイトーシスされ、処理され、最終的にT細胞に提示されて特定の免疫応答を誘導します。私たちの以前の研究では、モデル抗原であるオボアルブミン(OVA)を、N-グリカンG0を終結させる合成二分岐GlcNAcで機能化すると、DCへのターゲティングとそれに続く抗原の取り込みと提示が強化されることを示しました[21]。上記の現象は、DCの表面に発現するG0グリカンとエンドサイトーシスC型レクチン受容体の相互作用によって開始されると仮定しました。したがって、蛍光および多価G0-OVA金ナノクラスターは、以前の研究で適用された蛍光標識G0-OVAの代替となり、DC可視化の新しいツールとして使用できると考えています。さらに、強力なT細胞免疫応答の開始を可能にするDCのグリカン媒介ターゲティングを使用して、ワクチン候補の有効性を高めることができます[22、23、24]。この最初の研究では、ネオ糖タンパク質から始まる金ナノクラスターの合成と、レクチン凝集実験を使用したG0グリカンの機能性とアクセス可能性の評価を紹介します。最後に、樹状細胞のイメージングのための蛍光金ナノクラスターの可能性を示します。
結果と考察
非結合OVAタンパク質を使用した以前の報告に基づいて、G0グリカン(G0-OVA-AuNC)で装飾された卵白アルブミン金ナノクラスターの合成を最適化しました[25、26]。タンパク質で保護されたAuNCは、金のテトラクロロ金(III)酸(HAuCl 4 。 3H 2 O)タンパク質溶液に、続いて水酸化ナトリウムの1M水溶液に。反応pHを上げると、OVAに存在するトリプトファンおよびチロシン残基の還元電位が高まります(図1a)[14]。タンパク質足場は、還元試薬と安定化試薬の両方として機能し、タンパク質構造内に小さな金のクラスターを閉じ込め、環境から隔離します。蛍光OVA-AuNCを合成するための以前のプロトコルでは、非常に高濃度のOVA溶液(最大65 mg mL -1 )が必要でした。 )[25]。 AuNCの調製のための貴重なOVAネオ複合糖質の使用を制限するために、OVA-AuNCを効率的に生成できる非結合OVAの最小量を決定しました。 15 mg mL -1 のOVA濃度であることがわかりました 強い蛍光発光を持つクラスターを生成するのに十分でした(追加ファイル1:図S1)。 OVA-AuNCの形成は、マイクロ波照射によって大幅に加速され、反応時間が18時間から6分に短縮されました[25]。さらに、マイクロ波照射は溶液に均一な加熱を提供し、均一で単分散のクラスターの形成を促進しました[1]。この方法で調製されたOVA-AuNCは、淡褐色を示し、UV光照射下で強い赤色の蛍光を発します(図1b)。 OVA-AuNCのUV-Visスペクトルは、局在表面プラズモン共鳴バンドに対応する吸光度を欠いており、透過型電子顕微鏡(TEM)によってさらに確認された5nmを超える金ナノ粒子が存在しないことを示唆しています[27]。 1.9±0.7nmの範囲でOVA-AuNCsゴールドコアの平均直径を測定しました(追加ファイル1:図S2)。最後に、OVA-AuNCの平均流体力学的サイズ8.7nm±2.5nmが動的光散乱によって割り当てられました(追加ファイル1:図S2)。 OVA(直径6.5〜7 nm [28])と同様のサイズ範囲は、金のコアごとに1つのタンパク質分子が存在することを示しています。アガロースゲル電気泳動によるOVA-AuNCのさらなる分析は、正電極に対するOVAおよびOVA-AuNCの同様の移動度を示し、両方の種の同様のサイズと中性pHでのそれらの負電荷をさらに確認します(図1c)。 OVA-AuNCの蛍光発光スペクトルは、350nmで励起するとλ670nmに最大発光ピークを示します(図1d)。 AuNCの励起スペクトルは広く、ストークスシフトは大きく(200 nm以上)、青色発光のAlexaFluor®405色素など、同様の励起波長を特徴とする別の蛍光プローブ(多重化)の存在下でのスペクトル多色検出アプリケーションに最適です。 [26]。 0.1 M NaOH(QY =≈92%)中のフルオレセインを参照標準として使用した場合、OVA-AuNCの量子収率(QY)は約4%と計算されました[29]。金コアの酸化状態は、X線光電子分光法(XPS)測定に基づいてAu(I)種とAu(0)種の混合物に割り当てられました(追加ファイル1:図S3)[11、30]。タンパク質の二次構造の円二色性(CD)の研究により、ランダムコイル構成が明らかになり、おそらく合成中の過酷なアルカリ条件が原因で、AuNCのOVAのネイティブフォールドが失われることが示唆されました(追加ファイル1:図S4)[31]。最後に、広範囲のpH(3〜11)内、およびin vitro細胞培養で最も一般的に使用される血清サプリメントであるウシ胎児血清(FBS)を含む溶液でも、OVA-AuNCの並外れた安定性を観察しました。 (追加ファイル1:図S5)。 OVA金ナノクラスターのこの機能は、invitroおよびinvivoバイオアッセイでの有用性を強調し、刺激的な新しいアプリケーションを開きます。たとえば、OVA-AuNCのpHに影響されない蛍光発光により、わずかに酸性のpHを特徴とするエンドソームコンパートメント内でも信号を失うことなく、細胞内で効率的な粒子追跡が可能になります[32、33]。逆に、これらの条件下では、これらの色素の最大蛍光発光が塩基性pHで達成されるため、特定のフルオレセインコンジュゲートの使用が損なわれる可能性があります。 OVA-AuNCは、水と細胞増殖培地の両方で優れた溶解性も示しました。 OVA-AuNCの完全な可溶化は、水と細胞増殖培地の両方で40 mg / mLの濃度まで観察されました。 (追加ファイル1:図S6)。さまざまなOVA-AuNC希釈液での蛍光発光スペクトルを測定し、37°Cで24時間までのインキュベーションで安定していることを確認しました。
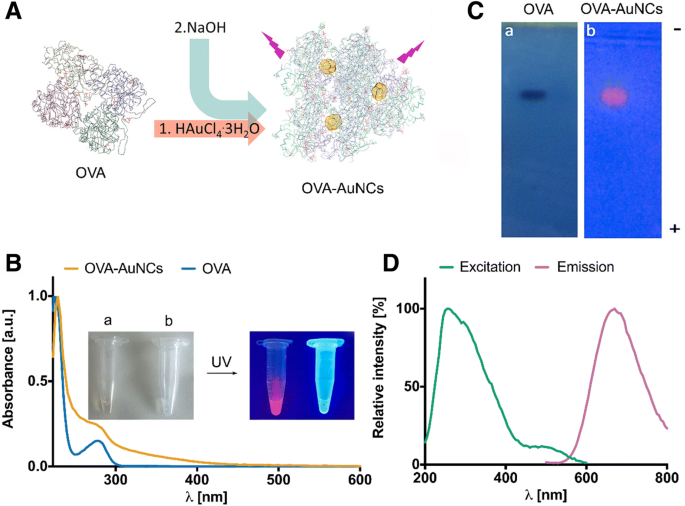
a OVA-AuNCs合成の概略図。 b OVA(青い線)およびOVA-AuNC(オレンジ色の線)のUV-可視スペクトル。挿入:OVA-AuNCの画像( a )およびOVA( b )可視光照明下(左)およびUV光照明下(365 nm)(右)。 c OVA(クマシーブルーG-250で染色)およびOVA-AuNC(UV光照射下)のアガロースゲル電気泳動。 d OVA-AuNCの蛍光励起(緑色の線)および発光(ピンク色の線)スペクトル
G0-OVAネオ糖タンパク質で保護されたAuNCの合成のために、最初にG0機能化OVAネオ糖タンパク質を調製しました。 C5アミノリンカーを備えたバイアンテナリーN-グリカンG0は、前述のように合成され[34]、OVAとの結合は、架橋試薬としてジスクシンイミジルスベレートエステル(DSS)を使用して達成されました(図2a)[21、35]。簡単に説明すると、結合は2段階の反応で実行されます。N-グリカンG0は13倍過剰のDSSリンカーで官能化され、OVA上の遊離アミノ基に結合します。糖鎖/タンパク質の比率を制御することにより、OVA置換の程度を効果的に調整することができました(SIを参照)。ネオ糖タンパク質の形成が成功したことは、SDS-PAGEゲル上に電気泳動的に遅い移動バンドが拡散することで確認されました(図2b)。一方、導入されたグリカンの平均数は、MALDI-TOF質量分析によってさらに決定されました[21](図2b)。 。2c)。この戦略に従って、タンパク質あたり2〜3、3〜4、および5〜6コピーのN型糖鎖G0を表示するOVAネオ複合糖質を作成しました。
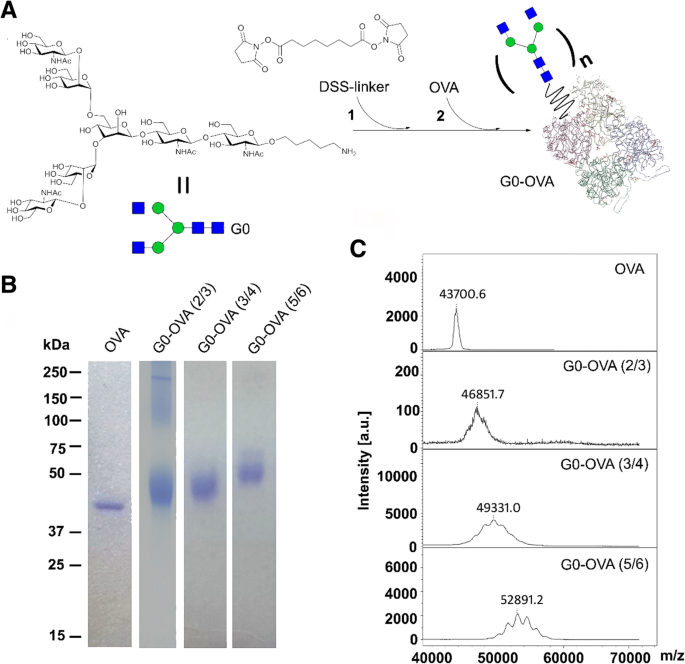
a G0-OVAネオ複合糖質の合成( n =原子価)。 b 異なる原子価のN-グリカンG0を含むOVAおよびG0-OVAネオ糖タンパク質のSDS-PAGEゲル電気泳動。 c 異なる原子価のN-グリカンG0を含むOVAおよびG0-OVAネオ糖タンパク質のMALDI-TOF質量スペクトル
次に、以前に最適化された条件下でAuNCの調製に合成ネオ糖タンパク質を使用し(図3a)、クラスターの物理的および光学的特性に対するグリカン官能基化とその原子価の影響を調査しました。図3に示すように、G0-OVA-AuNCの吸収UV-Visと蛍光発光はOVA-AuNCと同一であり、278 nmで特徴的な吸光度を示し、670 nm付近で最大の赤色蛍光発光を示しました(図3b、c)。
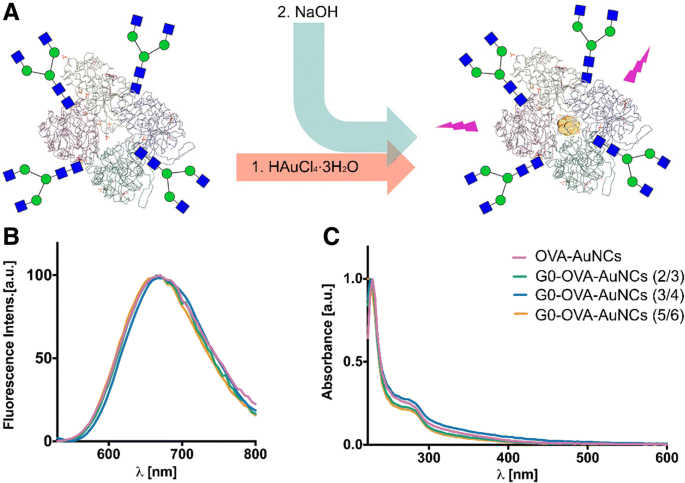
a G0グリカン由来のOVA-AuNCs合成の概略図。 b G0-OVA-AuNC(青、オレンジ、緑の線)とOVA-AuNC(ピンクの線)の蛍光発光スペクトル。 c G0-OVA-AuNC(青、オレンジ、緑の線)とOVA-AuNC(ピンクの線)のUV-可視スペクトル
さらに、TEM測定により、G0-OVA-AuNCコアの平均直径は1.6±0.5 nm(図4)であり、OVA-AuNCのサイズ(1.9±0.7 nm)に匹敵します。したがって、我々の結果に基づいて、リジン残基またはN末端を介してOVAに結合したグリカンは、糖タンパク質の全体的な還元能およびテストされた原子価でのクラスターのさらなる形成に影響を与えないと考えます[36]。さらに、糖は、タンパク質のシステイン基と金のコアの間のクラスター安定化Au(I)チオレートポリマー[37、38]の形成を妨げないようです。これは、多数(5〜6コピー)存在する場合でも同様です。
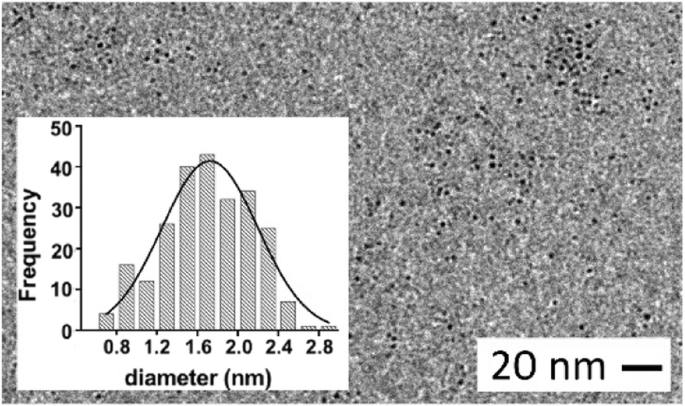
G0-OVA-AuNCのTEM画像(3/4)。挿入:G0-OVA-AuNCs(3/4)の金コア直径のサイズ分布
最後に、OVA-AuNCと同様に、G0-OVA-AuNC(3/4)タンパク質の二次構造がCDによって割り当てられ、ランダムコイル構成が明らかになりました(追加ファイル1:図S4)。ただし、AuNCsのタンパク質足場にグリカンを付着させることにより、炭水化物結合タンパク質との相互作用を可能にするターゲティング分子を導入すると同時に、OVAタンパク質は担体としてのみ残り、コンフォメーション変化によってレクチン認識が変化することはありません。システム全体。マウスにおける抗体の産生[39]およびT細胞の活性化における変性OVAの抗原能力が報告されています。 T細胞の活性化は、天然および変性OVAの両方で提示される特定のアミノ酸配列の認識を介して媒介されるため、抗原の二次構造とは無関係です[40]。実際、抗原OVA 323-339ペプチドはOVAに対する主要な特異的T細胞応答を説明し、このペプチドは免疫応答を誘導するためにナノ粒子に使用されています[41]。
ネオ糖タンパク質で保護された金ナノクラスターに新たに導入されたグリカン鎖の機能を調査するために、凝集実験でさまざまな植物レクチンとの相互作用を調べました。 G0-OVA-AuNCとOVA-AuNCを、末端のGlcNAc部分に特異的なBandeiraea simplicifoliaレクチン-II(BSL-II)と Solanum tuberosum の2つの異なる植物レクチンとインキュベートしました。 レクチン(STL)、N型糖鎖に存在するコアキトビオースを認識します[34、42]。さらに、G0-OVA-AuNCとレクチンの間の非特異的な相互作用を破棄するために、 Aleuria aurantia L-フコースに特異的なレクチン(AAL)を対照として含めた。 G0-OVA-AuNCs(5/6)の溶液に、BSL-II、STL、およびAALレクチンの濃度を上げていきました。暗所でのインキュベーション後、溶液を遠心分離し、上澄みの蛍光を測定した。図5に示すように、BSL-IIおよびSTLとのインキュベーション後、濃度依存的に蛍光強度の低下を観察できましたが(図5a〜d)、AALの存在下では蛍光強度は変化しませんでした(図5a〜d)。 。5e)。 G0-OVA-AuNCs(5/6)をSTLとインキュベートした後、UVランプ照射下で目に見える沈殿物が観察されましたが、AALとのインキュベーションでは沈殿物は現れませんでした(図5f)。レクチンインキュベーション後の初期蛍光値(F0)と最終値(F)の関係は、レクチン濃度の増加に対して表されました(図5b、d)。 STLとG0-OVA-AuNC(5/6)との相互作用は、0〜7.5μMの直線性を示し、検出限界は、式SD * 3 / S(SDは検量線の標準偏差で
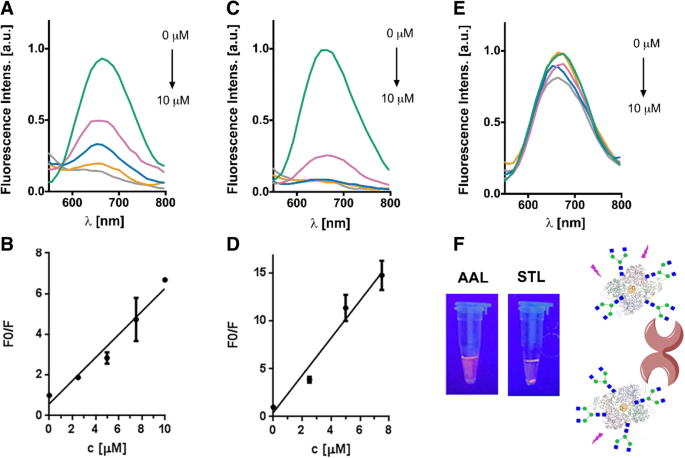
G0-OVA-AuNCのレクチン凝集アッセイ(5/6)。 a BSL-IIとのインキュベーション後のOVA-G0(5/6)-AuNCs溶液の上清の代表的な蛍光スペクトル。 b BSL-IIの濃度の増加に対するF0 / Fの表現。 c STLとのインキュベーション後のOVA-G0(5/6)-AuNCs溶液の上清の代表的な蛍光スペクトル。 d STLの濃度の増加に対するF0 / Fの表現。 e AALとのインキュベーション後のOVA-G0(5/6)-AuNCの上清の代表的な蛍光スペクトル。 f G0-OVA-AuNC(5/6)とAALおよびSTLの、UV光照射(365 nm)下でのインキュベーションに対応する画像。 STLとのインキュベーション後、目に見える沈殿物が形成されました。右側:G0-OVA-AuNCと植物レクチン間の結合の概略図
また、炭水化物タンパク質相互作用における培地成分の考えられる影響を識別するために、細胞増殖培地の存在下でのG0-OVA-AuNC(5/6)とSTLの相互作用を研究しました。 G0-OVA-AuNC(5/6)は、ウシ胎児血清を添加した完全Iscoveの改良ダルベッコ培地(IMDM)に溶解し、STLの量を増やしながら培養しました。得られた溶液をアガロースゲル電気泳動で分析した。 (追加ファイル1:図S8)。 G0-OVA-AuNCs(5/6)の電気泳動移動度は、水中と複雑な媒体の両方で維持されます。それにもかかわらず、増加する量のSTLの存在下では、G0-OVA-AuNC(5/6)の負の極への用量依存的な変位があり、複雑な細胞培地の存在下でもタンパク質炭水化物相互作用を強調しています。これは、タンパク質炭水化物とG0-OVA-AuNC(5/6)との相互作用が、培地成分の存在によって妨げられていないことを示しています。
蛍光イメージングのクラスターの可能性と、G0-OVAネオ糖タンパク質によるDCターゲティングでの以前の成功[21]により、invitroでのマウスDCによる取り込みにおけるG0-OVA-AuNCの研究が促進されました。共焦点蛍光顕微鏡を使用して、自己蛍光G0-OVA-AuNC(3/4)の内部移行を視覚化しました。脾臓DCは、C57BL / 6JマウスとCD11c + から分離されました。 集団は磁気活性化セルソーティング(MACS)(追加ファイル1:図S9)によって精製され、ポリ-d-リジンでコーティングされたガラスカバースリップに一晩播種されました。概念実証として、精製されたDCをG0-OVA-AuNC(3/4)とインキュベートし、洗浄して未結合の物質を除去し、固定しました。 40分間のインキュベーション後、共焦点蛍光顕微鏡画像を取得し(図6)、G0-OVA-AuNC(3/4)とインキュベートした樹状細胞の強い蛍光を観察し、効果的な内在化(図6a)と蛍光の欠如を示しました。ネガティブコントロール(刺激されていないCD11 + 細胞;追加ファイル1:図S9)。ナノクラスターの内部移行は、 Z に沿って撮影された単一のDC画像を積み重ねることによってさらに確認されました。 -軸(zスタック)および単一DCの3D画像を再構築して、細胞内からの蛍光発光を視覚化します(図6bおよび追加ファイル1:図S10)。
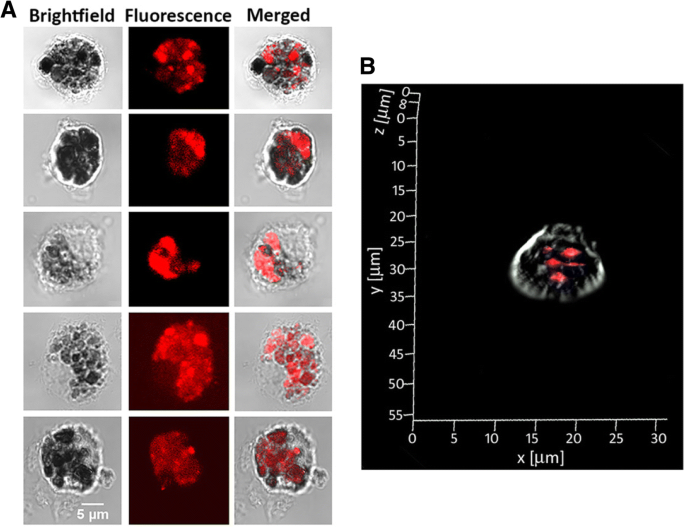
共焦点顕微鏡で測定したマウスDCによるG0-OVA-AuNC(3/4)の取り込み。 a CD11c + の代表的な画像 G0-OVA-AuNCs(3/4)とのインキュベーション後の細胞。 b 細胞内のG0-OVA-AuNCの取り込みを表す単一の樹状細胞のZスタック画像
要約すると、凝集実験およびDC取り込みアッセイで使用されているネオ糖タンパク質で保護された金ナノクラスターを準備し、特性を明らかにしました。特定の植物レクチンセンシングの例では、炭水化物-タンパク質相互作用の分析のためのG0-OVA-AuNCの有用性を示しました。また、蛍光顕微鏡による樹状細胞のinvitroイメージングを示しました。ただし、最初にDCによるナノクラスターの取り込みにおけるグリカンの役割をよりよく理解するために(たとえば、細胞内コンパートメント内のナノクラスターの局在を追跡することによって)、次にそれらの生体適合性および細胞適合性を研究するために、さらなる実験を行う必要があります。 G0グリカンのターゲティング特性と、非結合OVAと比較したDCによるG0-OVA取り込みの増加を説明した以前の結果に基づいて、G0-OVA-AuNCは蛍光標識されたネオ糖タンパク質の魅力的な代替品になる可能性があると考えています。
結論
結論として、この最初の研究では、ネオ糖タンパク質で保護された金ナノクラスターの最初の合成と、非複合タンパク質クラスターと比較したそれらの物理的および光学的特性の評価を提示します。植物レクチンとの特異的相互作用により、凝集アッセイにおけるAuNC上の糖G0のアクセス可能性を確認しました。これにより、レクチンの効果的な架橋活性のための最小グリカン密度の重要性がさらに強調されました。概念実証として、モデルマウスDCのイメージングにおけるG0-OVA-AuNCの適合性も実証しました。
自己蛍光性のネオ糖タンパク質で機能化されたAuNCは、炭水化物とタンパク質の相互作用の分析と応用を可能にする魅力的なツールになる可能性があると考えています。大きなストークスシフト、水溶性、pH安定性など、独自の物理的および化学的特性に基づいて、G0-OVA-AuNCは有機染料ラベルの代替となり、DCの視覚化、炭水化物を介した取り込み研究、および植物に使用できます。レクチンセンシング。最後に、糖鎖の付着にOVAのような免疫原性担体を使用することにより、ネオ糖タンパク質で機能化された金ナノクラスターは、抗原の取り込み、処理、および提示の詳細な研究を可能にするだけでなく、将来的には、蛍光治療薬またはアジュバント分子。
メソッド/実験
資料
すべての溶液は、Diamond UV浄水システム(Branstead International、IA、マドリード、スペイン)からのナノ純水(18MΩcm)で調製されました。 AuNCの調製に使用されたすべてのガラス器具は、事前に王水(HNO 3 )で洗浄されていました。 :HCl、1:3、 v / v )そしてナノ純水で十分にすすいだ。塩化金(III)三水和物、水酸化ナトリウム、およびトリエチルアミンはSigmaAldrichから購入しました。 LPSを含まない卵白アルブミンはHyglos(Bernried、Germany)から購入しました。スベリン酸ジスクシンイミジル(DSS)およびDMSOはThermo FisherScientificから購入しました。 N-グリカンG0は、前述のように化学的に合成されました[21]。
AuNCs合成。一般的な手順
OVAまたはネオ糖タンパク質の攪拌溶液(15 mg mL -1 )ナノ純水中、HAuCl 4 の0.1M水溶液 ・3H 2 O(4.2 mM、最終濃度)を滴下しました。得られた混合物を室温で5分間撹拌し、1 M NaOH水溶液(最終濃度150 mM)を滴下して混合物のpHを上げました。得られた溶液をBiotage®Initiatorマイクロ波リアクター内で100°Cで6分間インキュベートしました。 AuNCの透析は、Thermo FisherScientificのSlide-Z-lyserTM透析カセット(10 K MWCO)でナノ純水を介して実行されました。
合成されたOVAおよびネオ糖タンパク質で保護されたAuNCの蛍光発光スペクトルは、Nunc™96ウェルポリスチレンブラックプレートを使用して、Varioskan Flashmicroplateリーダー(Thermo Scientific)で、ScanItソフトウェアで動作するλ350nmの励起波長で測定されました。
OVA-AuNCとOVAタンパク質のアガロースゲル電気泳動は、0.75%アガロースゲルで80Vで30分間行いました。 OVAで保護されたAuNCの可視化は、UV光照射(365 nm)下で行われ、OVAタンパク質はクマシーブルーG-250で染色されました。
TEMイメージングは、加速電圧200kVのJOELJEM-2100F電界放出TEMで実施されました。 AuNCsソリューションは、極薄カーボンサポート(Ted Pella、レディング、米国)でコーティングされた銅TEMグリッドにドロップキャストされました。金ナノクラスターの平均直径は、ImageJ(Java)を使用して定量化されました。
G0-OVA-AuNC(5/6)と植物レクチンのインキュベーション
10マイクロリットルのG0-OVA-AuNC(5/6)[0.2 mg mL -1 ] TSMバッファー(20 mM Tris・HCl、150 mM NaCl、2 mM CaCl 2 、2 mM MgCl 2 、pH =7.4)をNunc™384ウェルポリスチレンブラックプレートに配置しました。続いて、10μLの Aleuria aurantia レクチン(AAL)、 Solanum tuberosum TSMバッファー中のレクチン(STL)およびBandeiraea simplicifoliaレクチン-II(BSL-II)を添加すると、最終タンパク質濃度は0、2.5、5、7.5、10μMになりました。対応する溶液を、穏やかに振とうしながら暗所で一晩インキュベートした。タンパク質溶液を11,000gで1時間遠心分離し、上澄みの蛍光発光スペクトルを、励起波長λ350nmのVarioskanFlashマイクロプレートリーダー(Thermo Scientific)で記録しました。 AuNCs-レクチン複合体の形成時の沈殿を観察するために、対照実験を行った。次に、10μLのG0-OVA-AuNC(5/6)[0.2 mg mL -1 ]を10μLのAALおよびSTL [40μM]とともに1時間インキュベートした後、11,000 gで遠心分離しました(1時間)。画像はUV光照射(365 nm)で撮影されました。
マウス脾臓樹状細胞の分離
すべての実験で使用されたマウス樹状細胞は、チャールズリバー(CIC biomaGUNE、サンセバスチャン、スペイン)によって飼育されたC57BL / 6Jマウスから分離されました。動物実験プロトコルは、CIC biomaGUNEの動物倫理委員会によって承認され、動物倫理と福祉に関する欧州連合のARRIVEガイドラインと指令に従って実施されました。マウスを従来の飼育条件(22±2°C、55±10%湿度、12時間の昼/夜サイクル)で飼育し、標準的な食餌を自由に与えました。 マウスを100%O 2 中の2.5%イソフルランで麻酔した 頸椎脱臼により安楽死させた。
脾臓はC57BL / 6Jマウス( n =3、女性、27〜28週齢)。脾臓細胞を分離するために、脾臓を2 mM l-グルタミン、100 U / mLペニシリン、100μg/ mLストレプトマイシン、および10%ウシ胎児血清(FCS; PAN Biotech)を添加したIscoveの改良ダルベッコ培地(IMDM)でフラッシュしました。細胞懸濁液を低温に保ち、40μmのセルストレーナーでろ過して細胞凝集体を除去しました。遠心分離(300 g、5分、4°C)後、細胞ペレットを5 mLの新たに調製した赤血球溶解バッファー(10%100 mM Tris pH 7.5 + 90%160 mM NH 4 )に再懸濁しました。 Cl)、穏やかに混合し、RTで2分間インキュベートします。細胞を完全なIMDM培地で2回洗浄し、遠心分離してからMACSバッファー(PBS、0.5%BSA、2 mM EDTA)に再懸濁しました。樹状細胞(CD11c + 細胞)は、CD11c + を使用した磁気活性化セルソーティングにより、C57BL / 6マウス脾臓細胞の懸濁液から分離されました。 マイクロビーズ(ミルテニー)。磁性マイクロビーズとともにインキュベートされた細胞は、磁場に置かれたMACSカラムにロードされました。 CD11c + を残したまま、結合していない細胞がカラムを通過しました 細胞をMACSバッファーで洗浄し、カラムから溶出しました。 DC純度を上げるには、CD11c + 細胞精製を繰り返した。細胞懸濁液を遠心分離し、IMDM完全培地に再懸濁し、カウントしました。
DCによるG0-OVA-AuNC(3/4)の取り込み
CD11c + cells from C57BL/6J mice (2 × 10 6 cells) were seeded on poly-d-lysine-coated glass cover-slips overnight. G0-OVA-AuNCs (3/4) were added to cells (150 μg mL −1 ) and incubated for 40 min at 37 °C. After incubation, cells were carefully washed with cooled PBS and fixed with 3% paraformaldehyde at RT for 20 min. After washing with PBS and water, the cover-slips were mounted on slides using Vectashield® mounting medium. Fluorescent images were taken using the Zeiss LSM 510 laser scanning confocal microscope (Carl Zeiss) equipped with a UV laser (365 nm) and × 63 oil immersion objective.
略語
- AAL:
-
Aleuria aurantia lectin
- AuNCs:
-
Gold nanoclusters
- BSA:
-
Bovine serum albumin
- BSL-II:
-
Bandeiraea simplicifolia lectin-II
- CD:
-
Circular dichroism
- CLRs:
-
C-type lectin receptors
- DCs:
-
Dendritic cells
- DMSO:
-
Dimethyl sulfoxide
- DSS:
-
Disuccinimidyl suberate ester
- FBS:
-
Fetal bovine serum
- G0:
-
Biantennary N-glycan
- G0-OVA-AuNCs:
-
Neoglycoprotein functionalized gold nanoclusters
- HAuCl4 . 3 H 2 O:
-
Gold tetrachloroauric (III) acid
- IMDM:
-
Iscove’s modified Dulbecco’s medium
- IR:
-
赤外線
- LOD:
-
Limit of detection
- MACS:
-
Magnetic-activated cell separation
- OVA:
-
Ovalbumin
- QY:
-
Quantum yield
- STL:
-
Solanum tuberosum lectin
- TEM:
-
透過型電子顕微鏡
- XPS:
-
X線光電子分光法
ナノマテリアル
- ビジョンとイメージングの最新アプリケーションの調査
- シクロカーボンの作成とイメージング
- 改善された診断および治療用途のための多機能金ナノ粒子:レビュー
- 合成および生物医学的応用のための蛍光ナノ材料の進歩と挑戦
- レーザーパルスとスパッタリング技術による導電性シリカナノファイバー/金ナノ粒子複合材料の合成
- 1D混合二元酸化物CeO2-LaOx担持金触媒の合成とCO酸化活性
- 光熱治療および光音響イメージングのためのポリピロール被覆鉄-白金ナノ粒子の合成およびinvitro性能
- ポリ(3,4-エチレンジオキシチオフェン)/金/グラフェン複合材料の固体加熱合成とその亜硝酸塩およびヨウ素酸塩のアンペロメトリー定量への応用
- 6-メルカプトプリンとニューロン透過性ペプチドで修飾された金ナノ粒子によるSH-SY5Y細胞増殖の促進
- 金および銀ナノ粒子のグリーン合成のためのPlatycodiRadix(Platycodon grandiflorum)からのPlatycodonサポニン
- 磁性金ナノ粒子標識ヘパラナーゼモノクローナル抗体とその後の腫瘍磁気共鳴画像法への応用