


ナノ粒子の細胞取り込みと細胞内輸送への洞察
要約
ナノ粒子科学は、さまざまな科学分野の展望を急速に変化させ、新しい技術プラットフォームを定義しています。これはおそらく、ナノ粒子が多くの病気の治療と診断のためのツールとして使用されてきたナノ医療の分野でさらに明白です。しかし、多大な利益がもたらされたにもかかわらず、この技術の一般的な落とし穴は、人体への潜在的な短期的および長期的な影響です。これらの問題を理解するために、多くの科学的研究が行われてきました。このレビューは、これらの研究のいくつかとその結果に光を当てようとしています。このレビューで検討されたトピックには、ナノ粒子のさまざまな可能な取り込み経路と細胞内輸送経路が含まれます。さらに、ナノ粒子の取り込みメカニズムと生物学的機能を決定する際の、サイズ、形状、電荷、表面化学などのナノ粒子の物理化学的特性の影響についても取り上げます。
はじめに
ナノ粒子(NP)は、その独自の特性と驚異的な適用性により、考えられるほぼすべての分野で現在最先端の研究の最前線にあるナノ材料のサブカテゴリです[1,2,3,4]。 RNCOSによる「GlobalNPMarket Outlook 2020」というタイトルの技術市場調査レポートでは、NPの市場は2015年から2020年の間に16%の複合年間成長率(CAGR)で成長すると報告されました。 NPテクノロジーは、急速に急成長しているアプリケーションのレパートリーにより、生物医学およびバイオテクノロジーの分野で独自のニッチを発見しました[5、6]。たとえば、NPは、薬物および遺伝子送達[7、8]、病原体のバイオ検出[9]、タンパク質の検出[10]、組織工学[11、12]、腫瘍のイメージングとターゲティング[13]、腫瘍の破壊に適用されています。温熱療法[14]およびMRI造影剤増強[15]。
サイズが小さいため、NPは細胞に容易に侵入し、細胞、組織、臓器を横切って移動することができます。 NPは、生物学的障壁を通過して細胞に入り、機能を発揮できるため、生物医学的用途で広く使用されています。ただし、両刃の剣のように、NPの潜在的なリスク(つまり、悪影響)もこの機能から発生します[16、17]。それらの「小さい」サイズにもかかわらず、極性分子としてのNPは細胞膜(CM)を通って拡散することができません。 CMはほとんどの場合、小さな非極性分子に対して透過性があるため、NPはエンドサイトーシス経路を使用して細胞に侵入します[18、19]。 NPが細胞に入る方法は、NPの生物医学的機能、生体内分布、および毒性を決定する上で重要な要素です。ナノメディシンでは、細胞へのNPの安全な侵入は、高い治療効果を得るための重要なステップです。さらに、NPの細胞内輸送と運命は、これらのキャリアが特定の細胞内コンパートメントを標的とし、造影剤、遺伝子、薬物などの特定の生体分子を送達することを目的としていることを考えると、NPの成功に不可欠なプロセスです[18、20、21、22 ]。さらに重要なことに、NPによる細胞毒性の誘導は、その侵入経路と細胞内局在によって決定されます。したがって、NPの細胞取り込みと細胞内輸送を理解することは、安全で効率的なナノメディシンを設計する上で非常に重要です[23]。
NPの細胞取り込み、ターゲティング、細胞内輸送は、サイズ、形状、表面特性などのNPの物理化学的特性を調整することで最適化できます[24]。したがって、細胞の取り込みに関与する根本的なメカニズムの知識は、NPの運命とその毒性を評価するために重要です。このレビューは、NPのさまざまな可能な取り込み経路とその細胞内輸送経路を強調しています。さらに、サイズ、形状、電荷、表面化学などのNPの物理化学的特性が、細胞による内在化に及ぼす影響についても取り上げます。細胞取り込みメカニズムに関連するNPの物理化学的特性を理解することで、周囲の健康な組織への毒性作用を最小限に抑えながら、制御された方法で標的作用部位に薬物ペイロードを送達するなど、生物医学的用途に不可欠な機能的NPを設計できます。と臓器。
NPの細胞取り込み経路
原形質膜としても知られるCMは、細胞外液から細胞内を切り離すことによって細胞質を囲みます。 CMは、細胞内成分を保護し、細胞の恒常性を維持し、構造的サポートを与え、細胞の組成を保持するため、非常に重要です[25、26、27、28、29]。 CMは、タンパク質が埋め込まれた二重層に配置されたリン脂質で構成されています。これらのリン脂質二重層は、親水性のヘッドと疎水性のテールを備えており、小さな生体分子の侵入を可能にします。より具体的には、CMは細胞への物質の通過を制御する選択的に透過性のバリアです[30、31]。 CMは、主に受動輸送と能動輸送の2つのカテゴリーに分けられる物質を交換するために、さまざまなメカニズムを採用しています。酸素や二酸化炭素などのガス、ベンゼンなどの疎水性分子、水やエタノールなどの非荷電分子は、高濃度から低濃度の領域から膜全体に拡散します。濃度勾配に沿っており、エネルギーの助けなしに発生するこの種の輸送は、受動輸送と呼ばれます。対照的に、能動輸送は、アデノシン三リン酸(ATP)によって提供されるエネルギーを使用することによって濃度勾配に対して発生します[32、33、34、35、36]。
疎水性原形質膜を通過できない極性または荷電生体分子は、エンドサイトーシスと呼ばれる能動輸送の形態によって内在化されます。この過程で、細胞はCMの陥入によって細胞外液内の物質を飲み込み、細胞内で芽を出し、エンドソームと呼ばれる膜結合小胞を形成します[37]。エンドサイトーシスは、基本的に、食作用と飲作用の2つの主要なカテゴリーに分類できます。食作用(細胞を食べる)は、食細胞(単球、マクロファージ、好中球など)と呼ばれる特殊な哺乳類細胞によって破片、細菌、またはその他の大きなサイズの溶質を取り込むプロセスです[38、39]。
食作用に不可欠なのはオプソニン作用と呼ばれるプロセスであり、免疫グロブリンや補体タンパク質などのオプソニンが標的物質をコーティングして、食細胞の存在を誘発し、食作用活性を初期化します[40]。食細胞が標的物質を摂取し始めると、同時に、摂取された物質が食細胞内に区画化されるファゴソームと呼ばれる膜結合小胞の形成を刺激します。このプロセスの後半では、ファゴソームがリソソームと融合し、リソソーム内腔に含まれる加水分解酵素によって酸性pHで物質が消化されます[41、42、43]。
すべての細胞タイプで、ナノメートルの範囲内の小さな粒子は飲作用によって内在化されます[44]。飲作用では、「細胞飲用」原形質膜が陥入を形成して、溶解した分子を含む細胞外液の小さな液滴を取り込みます。飲作用は識別プロセスではなく、細胞の必要性に関係なく、ほとんどすべての細胞で継続的に発生します。つかまれた物質は、ピノソームと呼ばれる小さな小胞にピンチオフされ、リソソームと融合して内容物を加水分解または分解します[45、46]。食作用と飲作用は、それらのエンドサイトーシス小胞のサイズによって区別することができます。前者は250nmのサイズの大きなベシクルによる大きな粒子の取り込みを含み、後者は数ナノメートルから数百ナノメートルのサイズの小さなベシクルを介した流体の取り込みを含みます[42、47]。飲作用は、クラスリン介在性エンドサイトーシス、カベオラ介在性エンドサイトーシス、クラスリンおよびカベオラ非依存性エンドサイトーシス、およびマクロ飲作用に細分類できます[48、49]。
クラスリンを介したエンドサイトーシスは、特定の分子を細胞に内在化するための細胞侵入メカニズムです。この侵入経路は、細胞が低密度リポタンパク質受容体によるコレステロールやトランスフェリン受容体による鉄などの原形質膜成分や栄養素を取り込むのを助けます[50、51、52、53、54、55、56]。このプロセスでは、細胞外液中の特定のリガンドがCMの表面の受容体に結合し、リガンド-受容体複合体を形成します。このリガンド-受容体複合体は、クラスリンが豊富なCMの特殊な領域に移動し、クラスリンでコーティングされた小胞の形成によって飲み込まれます。細胞内に入ると、小胞の外側のクラスリンコーティングは、初期のエンドソームと融合する前に排出されます。初期のエンドソーム内の積荷は、最終的にはエンドリソソーム経路を介してリソソームに到達します[40、57、58、59、60]。各タイプのNPは、優先的に取り込み経路を介して細胞によって内在化されます。たとえば、ポリ(乳酸-co-グリコール酸)、D、L-ポリラクチド、ポリ(エチレングリコール-ラクチド)とシリカ(SiO 2 )ベースのナノ材料は、クラスリンを介したエンドサイトーシス経路によって内在化されます[61]。クマリンベースの固体脂質NPは、これらのNPの構造がCMに類似しているため、エネルギーに依存しない経路を介して細胞によって内在化されます。すべての脂質ベースのNPは、クラスリンを介したエンドサイトーシス経路を利用します[62]。ハーセプチンでコーティングされた金のNPは、膜ErbB2受容体による受容体を介したエンドサイトーシスを介して細胞に侵入します[63]。
カベオラを介したエンドサイトーシスは、カベオラ(小さな洞窟)と呼ばれるフラスコ型の膜陥入を伴う細胞侵入の経路です。カベオラは、内皮細胞、上皮細胞、脂肪細胞、筋肉および線維芽細胞に存在します[64、65、66、67]。カベオラのサイズは通常50〜80 nmの範囲であり、膜タンパク質のカベオリン1で構成されており、フラスコ型の構造を与えます[68,69,70,71]。カベオラ依存性エンドサイトーシスは、細胞シグナル伝達と膜タンパク質、脂質、脂肪酸の調節に関与しています[61、64、67]。カベオラが原形質膜から分離されると、中性pHで存在するカベオソームと呼ばれる細胞区画と融合します。カベオソームはリソソームをバイパスすることができるため、加水分解酵素およびリソソーム分解から内容物を保護します。したがって、ウイルスや細菌などの病原体は、分解を防ぐためにこの侵入経路を使用します。カベオリン依存性メカニズムによって細胞内に取り込まれたカーゴはリソソームに到達しないため、この経路はナノメディシンで採用されています[54、72、73、74]。
クラスリンおよびカベオラに依存しないエンドサイトーシスは、クラスリンおよびカベオリンを奪われた細胞で発生します。この経路は、成長ホルモン、細胞外液、グリコシルホスファチジルイノシトール(GPI)結合タンパク質、およびインターロイキン-2によって利用されて細胞に入ります。たとえば、クラスリンおよびカベオラに依存しない経路を使用して細胞に入る葉酸[58、72、75、76、77、78、79]は、ドラッグデリバリーシステムおよび造影剤として使用されるNPおよびポリマーに結合します[53 、80、81]。マクロピノサイトーシスは、細胞がマクロピノソームと呼ばれる大きな小胞(0.5〜10μm)を形成することによって大量の細胞外液を取り込む一種の飲作用メカニズムです[82,83,84,85]。マクロ飲作用は、アポトーシスおよび壊死細胞、細菌およびウイルス、ならびに抗原提示を内在化するための経路です。この経路は、他のほとんどの経路では細胞に取り込まれないミクロンサイズのNPを内在化する可能性があります。マクロ飲作用は、脳微小血管内皮細胞を除くほとんどすべての細胞で発生する可能性があります[86,87,88,89]。 NPは、図1に示すように、これらのエンドサイトーシス経路の1つを介して細胞に入ります。
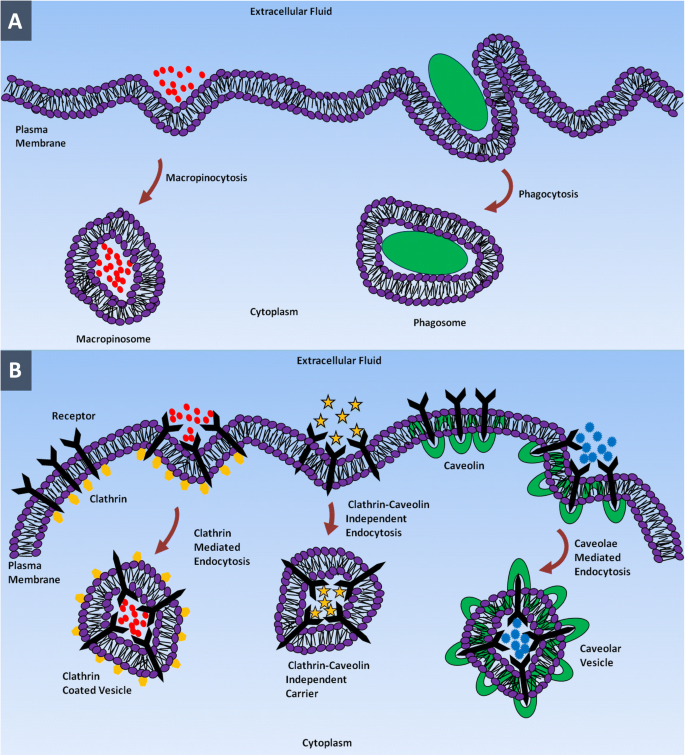
異なるエンドサイトーシス経路を使用した細胞へのNPの侵入。 a マクロ飲作用および食作用。 b クラスリン介在性エンドサイトーシス、クラスリン-カベオリン非依存性エンドサイトーシス、およびカベオラ介在性エンドサイトーシス
細胞取り込みに対するNPの物理化学的特性の影響
サイズ、形状、表面電荷、表面疎水性/親水性、表面機能化などのNPの物理化学的特性が細胞取り込みに及ぼす影響を研究することは、これらのパラメーターがNPの取り込みレベル、エンドサイトーシス経路、および細胞毒性に直接影響するため、非常に重要です。 [90、91]。 NPの細胞取り込みに影響を与える物理化学的要因を図2に示します。次のセクションでは、これらのパラメーターが細胞とNPの相互作用に与える影響について説明します。
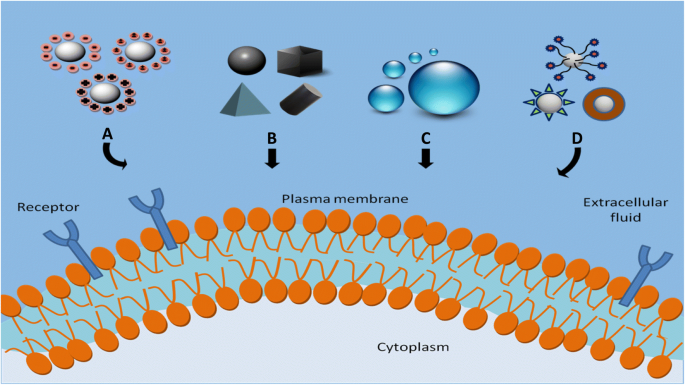
NPの細胞取り込みに影響を与える物理化学的要因。 a 表面電荷、 b 形、 c サイズと d 表面化学
サイズの影響
NPのサイズは、細胞取り込みの効率[92]と、生細胞に対する毒性の可能性[24]を決定する重要な要素です。さらに、NPのサイズは、取り込み経路の決定にも大きな役割を果たすことがわかりました。数から数百ナノメートルの範囲のサイズの小さなNPは、飲作用またはマクロ飲作用を介して細胞に入ります。 250 nm〜3μmのサイズ範囲のNPは、最適なin vitro食作用を示すことが示されていますが、120〜150 nmのサイズ範囲のNPは、クラスリンまたはカベオリンを介したエンドサイトーシス、およびNPの最大サイズを介して内在化されます。この経路を使用することは200nmであると報告されました[47、93]。カベオラを介した経路では、カベオラのサイズがより大きなNPの取り込みを妨げます[16、17]。特定のタイプのNPは、そのサイズに応じて複数のエンドサイトーシス経路を利用する場合があります。
いくつかの研究は、NPの細胞取り込みについて、NPがより効率的に内在化され、より高い取り込み速度を持つ50nmの最適サイズがあることを示しています。 NPの取り込みは、小さい粒子(約15〜30 nm)または大きい粒子(約70〜240 nm)で減少することが示されました[94,95,96,97,98,99]。さらに、30〜50 nmのサイズのNPは、CM受容体と効率的に相互作用し、その後、受容体を介したエンドサイトーシスを介して内在化されます[97]。 NPのドラッグデリバリーアプリケーションでは、主な関心事は、NPが細網内皮系によって排除されるのを防ぎ、血中の循環時間を延長して、標的でのバイオアベイラビリティを高めることです。この点で、NPのサイズを大きくすると、クリアランス率が高くなります[100,101,102,103,104,105]。したがって、細胞への取り込みにおけるNPサイズの役割を理解することは、医療用途向けの効果的で安全なNPを設計するために重要です。
さまざまな研究がNPのサイズと取り込み経路の関係を調査しましたが、明らかにされた結果は常に一貫性がありませんでした[93、106、107、108、109]。これらの矛盾は、サイズを制御するプロセス中にNPの他のパラメーターを制御することの複雑さに関連している可能性があります。それに加えて、合成後に測定されたNPのサイズは、凝集および凝集のためにinvitroおよびinvivoの研究中に変化する可能性があり、これが細胞内在化経路に影響を与える可能性があります[110、111]。非食細胞B16細胞の細胞取り込み経路に対する粒子サイズの影響は、50〜1000nmの範囲のさまざまなサイズの蛍光ラテックスビーズを使用して調査されました[93]。結果は、これらのビーズの内在化メカニズムが粒子サイズに大きく依存していることを示しています。特に、200 nm以下のサイズのビーズはクラスリンでコーティングされたピットに取り込まれましたが、より大きなビーズはカベオラを介したエンドサイトーシスによって内在化されました。 Laiと共同研究者[16]は、25 nm未満のサイズの小さな高分子NPが、エンド/リソソーム経路の外側の非分解性小胞を介して細胞の核周辺領域に到達するための新しいメカニズムを採用していることを発見しました。この経路は、クラスリンおよびカベオラを介さず、コレステロールに依存しません。
SK-BR-3細胞によるハーセプチン-AuNPと結合した異なるサイズ(2〜100 nm)の金(Au)NPの取り込みは、サイズに依存することが示されました。最高の細胞内在化は、25〜50nmのサイズ範囲のNPで観察されました[63]。この侵入経路では、NPのサイズが、膜受容体の結合と活性化、およびタンパク質の最終的な発現の決定要因であることがわかりました。コロイド状AuNPのサイズと形状の変化が細胞内取り込みに及ぼす影響を評価しました[112]。球形および棒状の14、50、および74 nmサイズのAuNPを、HeLa細胞とインキュベートしました。 NPの取り込みはそのサイズと形状に強く依存し、50nmのサイズの粒子が最も高い取り込み速度を示したことがわかりました。さらに、球状のAuNPの取り込みは、同様のサイズの棒状のNPよりも500%多かった。シャンら[113]は、HeLa細胞による直径4、12、および17nmのAuNPのエンドサイトーシスのサイズ依存力を調査しました。結果は、取り込み力と非結合力の両方の値が、AuNPのサイズによって増加することを明らかにしました。 SiO 2 の取り込み A549細胞(肺上皮細胞)による異なるサイズ(50、100、300 nm)のNPは、フローサイトメトリー、蛍光、電子顕微鏡の組み合わせによって研究されています。これらの研究者は、SiO 2 の取り込みが NPはサイズによって減少しました[114]。
形状の影響
サイズに加えて、NPの形状は、NPの輸送だけでなく取り込み経路においても極めて重要な役割を果たします。 Chithrani etal。 [112]は、コロイド状AuNPの形状がHeLa細胞の取り込みに及ぼす影響を研究しました。その結果、球状のAuNPは棒状のAuNPよりも5倍高い取り込み率を示しました。別の研究では、同じ研究者が、3つの異なる細胞株での球状および棒状のトランスフェリン被覆AuNPの取り込みレベルを調査しました。 STO細胞、HeLa細胞およびSNB19細胞[94]。彼らは、球状のAuNPが、棒状のAuNPよりも高い速度ですべての細胞株に取り込まれることを観察しました。
インビボでの形状の効果を確立するために、Gengと共同研究者[115]は、齧歯動物における球体を伴う柔軟なフィラメントの輸送と輸送の違いを評価するためにフィロミセルを採用しました。その結果、フィロミセルは球状のフィロミセルの約10倍の循環を維持していることが明らかになりました。さらに、球体フィロミセルは、より長いフィラメントよりも容易に細胞によって内在化される。 Grattonと共同研究者[106]は、単分散ヒドロゲル粒子の形状がHeLa細胞への取り込みに及ぼす影響を実証しました。彼らは、棒状のNPが球、円柱、立方体と比較して最も高い内在化率を持っていることを発見しました。別の研究では、細胞取り込みに対するNPの形状の影響を、Caco-2細胞に円盤状、球形、および棒状のポリスチレン(PS)NPを使用して調査しました。結果は、棒状および円盤状のNPが球形のNPより2倍高く内在化されたことを示した。彼らは、NPの形状を考慮することにより、NPを介したドラッグデリバリーを進めることができると結論付けました[116]。
Xuと共同研究者[117]は、六角形のシート(横方向に50〜150 nm、横方向に10〜20 nm)などのさまざまな形態のフルオレセインイソチオシアネート(FITC)を使用して層状複水酸化物(LDH)NPを調製することにより、細胞への取り込みに対する形状の影響を研究しました。太い)およびロッド(幅30〜60 nm、長さ100〜200 nm)。すべての形態は、クラスリンを介したエンドサイトーシスを介して取り上げられました。 LDH-FITCナノスフェアは細胞質に保持されたが、LDH-FITCナノロッドは微小管によって核に向かって移動した。 Dasgupta etal。シミュレーションを適用して[118]、細胞取り込みにおけるNPの形状の役割を調べました。彼らは、ナノロッド型およびナノキューブ型のNPの膜ラッピングをシミュレートしました。棒状粒子の場合、ラッピング率が小さくて高い安定したエンドサイトーシス状態が見つかりました。アスペクト比の増加は、完全なラッピングには望ましくありませんでした。 NangiaとSureshkumar [119]は、高度な分子動力学シミュレーション技術を適用することにより、NPの転座率に対する形状の影響をコンピューター化しました。この研究の主な発見は、円錐形、立方体、棒、米、ピラミッド、球形のNPの転座率の有意な変動です。
表面電荷の影響
NPの細胞取り込みに影響を与えるもう1つの重要な要因は、表面電荷です。最近の10年間で、ナノ表面修飾がNPの表面電荷をカチオン性またはアニオン性のいずれかに設計するために採用されてきました[92]。負に帯電したCMは、正に帯電したNPの取り込みを強化します。特に、正に帯電したNPは、中性および負に帯電したNPよりも内部移行が高くなります[47、120]。ただし、正に帯電したNPの取り込みは、CMの完全性を破壊し、毒性の増加につながる可能性があります[121、122]。一般に、正に帯電したNPは細胞死を誘発します[123、124]。興味深いことに、中性に帯電したNPは、負に帯電したNPと比較して細胞への取り込みを低下させます[110、125、126、127]。さらに、負に帯電したNPの内部移行は膜のゲル化を引き起こし、正に帯電したNPはCMに流動性を引き起こします[128、129]。 NPの取り込み率に加えて、表面電荷も取り込みメカニズムに影響を与えます。より具体的には、正に帯電したNPは主にマクロ飲作用を介して細胞に内在化されますが、クラスリン/カベオラ非依存性エンドサイトーシスは負に帯電したNPの取り込みのメカニズムです[130]。 AuNPの表面が有機分子でコーティングされている場合、細胞の取り込み経路は異なります。たとえば、正に帯電したプレーンなAuNPは、マクロ飲作用とクラスリンおよびカベオリンを介したエンドサイトーシスを介して内在化されますが、負に帯電したポリエチレングリコール(PEG)でコーティングされたAuNPは、主にカベオリンおよび/またはクラスリンを介したエンドサイトーシスを介して内在化されます[131]。 / P>
LiとGu [132]は、分子動力学シミュレーションを使用して、荷電および中性NPとCMとの相互作用を研究しました。帯電したNPは、中性のNPと比較してCMへの接着性が優れていることがわかりました。さらに、NPの電荷密度を上げることにより、NPを膜で完全に包むことができます。別の研究グループは、分子動力学シミュレーションを使用して、カチオン性およびアニオン性のAuNPとCMとの相互作用を調査しました。その結果、AuNPの電荷密度が高くなると、AuNPの浸透によるCMの破壊が増加することが明らかになりました[133]。これらの発見は、AuNPの表面電荷密度を操作して、生物医学的用途で検討されているNPの本質的な特性である細胞毒性を最小限に抑えながら、その取り込みを最適化することにより、細胞とAuNP間の相互作用を制御する方法を示唆しています。
LiとMalmstadt [134]は、正および負に帯電したPS-NPと生体膜との相互作用を研究しました。その結果、カチオン性NPと膜のリン酸基との間の強力な静電相互作用により、NP-膜結合と膜表面張力が強化され、その結果、細孔が形成されることが示されました。 SK-BR-3細胞への正に帯電したAuNPの取り込み率は、負に帯電したAuNPよりも5倍高いと報告されました。これらの研究者はまた、正に帯電したAuNPが非エンドサイトーシス経路によって内在化され、負に帯電したAuNPがエンドサイトーシス経路を介して細胞に取り込まれることを調査しました[135]。
Hauck etal。 [107]は、HeLa細胞による18〜40 nmのサイズ範囲と+ 37 mV〜- 69 mVの範囲の表面電荷を持つ金ナノロッド(AuNR)の取り込みを調べました。結果は、AuNRのすべての濃度で、HeLa細胞への最高の内在化は+ 37 mVの表面電荷であり、最低の内在化は-69mVであったことを示しました。 Huhnと共同研究者[136]は、コロイド状AuNPと、3T3線維芽細胞、マウスC17.2神経前駆細胞、ヒト臍帯静脈内皮細胞などのさまざまな細胞株との電荷依存性相互作用を評価しました。結果は、すべての細胞株について、カチオン性AuNPがアニオン性対応物よりも高い取り込みを示したことを示した。彼らは、細胞の取り込みは電荷の兆候に大きく依存していると結論付けました。さらに、細胞毒性研究は、正に帯電したNPの取り込みが高い結果として、負に帯電したNPよりも高い毒性を示すことを示しました。
疎水性の影響
NPの疎水性は、CMとの相互作用の決定要因です[92、137]。いくつかの研究は、CMとの相互作用に対するNPの疎水性の影響を示しました。 Li etal。 [138]は、分子動力学シミュレーションを使用して、CMとの相互作用に対するNPの疎水性/親水性の影響を研究しました。結果は、疎水性NPがCMに包含を作成し、親水性NPがCMに吸着することがわかったことを明らかにしました。別の研究では、NP-細胞相互作用に対する疎水性の影響を調査するためにシミュレーションアプローチが適用されました。親水性のNPが包まれているのに対し、疎水性のNPは、膜に直接浸透することにより、二重層の内側の疎水性コア内に埋め込まれていることが観察されました[139]。
混合脂質/高分子膜とのQDNPの相互作用は、NPの疎水性表面を変更することによって評価されました。疎水性NPは膜の混合脂質/ポリマー単層のポリマードメイン内に位置するのに対し、親水性QDNPは単層に吸着して全体に広がり、空気/水界面での分子パッキングへの影響が大きいことを示しています[140]。 。疎水性および親水性リガンドが混合された官能化AuNPのリポソーム壁への組み込みが研究された。結果は、疎水性リガンドが二重層の疎水性コアと相互作用し、親水性リガンドが水溶液と相互作用することを示しました[141]。
表面改質の効果
NPの生物医学的応用において、NPの表面化学修飾は、毒性を低減し、安定性を高め、NPの細胞内在化を制御および調節するために利用される重要なステップであり、したがってそれらの生物学的運命です[142]。 NPの表面官能化は、主にPEG、負のカルボキシル(–COOH)基、ヒドロキシル(–OH)基などの中性官能基、および正のアミン(–NH2)基で構成されます。 (–NH2)の量が増えると、正の表面電荷が増加するため、細胞へのNPの取り込みが増加します[143、144、145、146]。同様に、–COOH官能基は、NPの負電荷を増加させ、それに応じてその取り込みを強化します[144]。
タオら[147]は、腫瘍を標的とするためのポリドーパミン官能化NP-アプタマーバイオコンジュゲートを設計しました。彼らは、機能化されたNPは、機能化されていないNPと比較してより優れたターゲティング効果を示し、機能化されたNPの細胞取り込み率が高く、治療効果が向上することを示しています。別の研究では、葉酸で機能化されたNPは、機能化されていないNPよりも子宮頸がん細胞の標的化において高い有効性を示しました[148]。 AuNPの毒性と細胞への取り込みに対する表面コーティングの影響は、Qiuとその同僚によって研究されました[90]。彼らは、ポリ(ジアリルジメチルアンモニウムクロリド)でコーティングされたAuNRが細胞による内在化においてより高い効率を示したため、表面コーティングが細胞取り込み速度を決定する重要な要因であることを明らかにしました。
未処理のポリスチレン(PS-NP)とアミノ官能化ポリスチレンNPの細胞への取り込みの違いは、Jiangとその同僚によって調査されました[149]。結果は、アミノ官能化ポリスチレンNPがPS-NPよりも高い取り込み率を示し、前者は主にクラスリン媒介経路を介して、後者はクラスリン非依存性エンドサイトーシスを介して内在化されたことを示しています。この顕著な違いは、NPとの細胞相互作用における表面化学修飾の重要な役割を浮き彫りにします。表面修飾フラーレン、C 60 ( C ( COOH ) 2 ) 2 NPは、主にエンドサイトーシスを介して、時間、温度、およびエネルギーに依存して細胞に内在化されました。クラスリンを介したエンドサイトーシスは、C 60 の内在化に好ましい経路であることがわかりました。 ( C ( COOH ) 2 ) 2 NP [150]。
弾力性の効果
NPの弾力性は、細胞による内在化に影響を与える内因性の要因です。 NPの弾性は、力が加えられたときの変化に対する抵抗によって説明できます。剛性、硬度、剛性は、NPの弾性を説明する際の同義語の一部です。 NPの弾性を測定するために使用されている測定指標はヤング率であり、測定単位はパスカル(Pa)です。この測定に基づくと、ヤング率の値が高いほどNPの弾性が高いことを示し、その逆も同様です。 NPでこの値を測定するために使用される分析デバイスまたは機器の例は、原子間力顕微鏡、レオメーター、およびナノインデンターです。弾性値が高いNPはハードNPと呼ばれ、その例として、ゴールドNP、量子ドット、磁気NPがあります。弾性値が低いNPはソフトNPと呼ばれ、これらの例としては、ヒドロゲル、リポソーム、生分解性ポリマーがあります。
細胞取り込みに関してNPのこのパラメーターに焦点を合わせた多くの研究は、より柔らかいNPと比較してより効率的により硬いNPを内在化する細胞の選好について報告しました[151、152]。明らかに、この観察結果は、NPを包むのに必要な変形エネルギーが内部移行プロセス全体で変化する場合でも、柔らかいNPと比較して硬いNPを包む際の膜による全体的なエネルギー消費が少ないことに起因します。さらに、粗視化分子動力学(CGMD)シミュレーションを使用して実施された、弾性が変化するNPの膜ラッピングの計算モデリングは、硬いNPと柔らかいNPの内部移行に伴う変形エネルギー変化に関する実験的観察と一致します[153]。 However, there are also other studies that have reported on softer NPs being internalized more efficiently than stiffer NPs [154, 155] and intermediate elastic NPs internalized more efficiently compared to either stiff or soft NPs [156]. Hence, tuning the elasticity of NPs for better cellular internalization could be a valuable tool in biomedical applications such as drug delivery. A potential application was demonstrated by Guo and coworkers, whereby accumulation of nanolipogels in tumour cells were enhanced primarily by controlling this parameter of NP [157].
Intracellular Trafficking of NPs
In the previous sections, different possible uptake pathways of NPs and the parameters that affect the efficacy of uptake has been discussed. Following uptake, the next crucial matter is the intracellular trafficking of NPs which determines its final destination within cellular compartments, its cytotoxicity and its therapeutic efficacy [158, 159]. After NPs are internalized by the cells, they will first encounter membrane-bound intracellular vesicles called early endosomes. Endosomes formed at the plasma membrane are categorized into three types; early endosomes, late endosomes and recycling endosomes [106, 160,161,162,163].
Early endosome ferries the cargo to the desired cellular destination. Part of the cargo is recycled to the plasma membrane via recycling endosomes. Early endosomes transform into late endosomes via maturation and differentiation process. The late endosomes will then integrate with lysosomes to form endolysosomal vesicles and hydrolytic enzymes contained within these vesicles degrade the trapped NPs [18, 164,165,166]. However, some NPs are able to escape this pathway and are released into the cytoplasm therefore bypassing the lysosomal degradation process [167,168,169]. Another intracellular degradation pathway which plays important role in the intracellular fate of NPs is an intracellular process called autophagy [170,171,172]. In this process, cytoplasmic contents will be surrounded by autophagosome and delivered to the lysosome to be broken down and recycled [173]. In addition, aggregated proteins and dysfunctional organelles are degraded by autophagy to maintain cellular homeostasis. It is necessary to consider this pathway since recent studies demonstrated that several NPs are capable of inducing autophagy [174,175,176,177,178].
The intracellular trafficking of Tat peptide-conjugated quantum dots (Tat-QDs) in live cells was studied by Ruan and co-workers [179]. Dynamic confocal imaging showed that Tat-QDs interacted with negatively charged CMs leading to its internalization by macropinocytosis. The QD containing vesicles were observed to be actively transported by molecular motors towards the perinuclear region known as the microtubule-organizing center (MTOC). Tat-QDs bind to cellular membrane structures such as filopodia and vesicle shedding results in releasing QD-containing vesicles from the tips of filopodia.
The uptake and intracellular fate of fluorescent carboxylated polystyrene particles (20 nm and 200 nm in diameter) were evaluated by applying it on hepatocyte [180]. It was found that the particles were internalized by hepatocytes in size, time and serum-dependent manner. The fate of the particles was studied and they were not observed in early endosomes or lysosomes, but only in the mitochondria of the hepatocyte. Particles accumulated inside bile canaliculi show that NPs can be eliminated within bile. A study on the uptake and intracellular fate of silver NPs into human mesenchymal stem cells demonstrated that they agglomerate in the perinuclear region [181]. It was observed by using fluorescent probes that particles are contained within endo-lysosomal structures but not in the cell nucleus, endoplasmic reticulum or Golgi complex. Confocal imaging of FITC conjugated titania nanotubes in mouse neural stem cells revealed that they have crossed the karyotheca entering the cell nucleus [182]. Single-walled carbon nanotubes were observed to enter the cytoplasm and localize in the cell nucleus leading to cell mortality [183]. Translocation of AuNRs towards the nucleus has also been reported [184].
結論
The application of NPs in the modern world is growing at an exponential rate as the scientific enterprise is looking for novel ways to address current problems. NPs can be found as active ingredients in many formulations intended for human consumption, from cosmetics to processed foods. As its application increases in consumer products, so does human exposure to NPs. Hence, more research should be carried out to understand its potential hazards to humans and other living beings. In this review, we have looked at the current knowledge on the effects of NPs at a cellular level. Some of the topics discussed include cellular pathways of NPs and the influences of physiochemical properties of NPs on the uptake rate and uptake mechanism.
略語
- ATP:
-
Adenosine triphosphate
- CAGR:
-
Compound annual growth rate
- CM:
-
Cell membrane
- FITC:
-
Fluorescein isothiocyanate
- GPI:
-
Glycosylphosphatidylinositol
- LDH:
-
Layered double hydroxide
- MTOC:
-
Microtubule-organizing center
- NP:
-
ナノ粒子
- PEG:
-
ポリエチレングリコール
ナノマテリアル
- 癌治療のためのナノ粒子:現在の進歩と課題
- 生体適合性FePO4ナノ粒子:ドラッグデリバリー、RNA安定化、および機能的活性
- コバルトをドープしたFeMn2O4スピネルナノ粒子の調製と磁気特性
- Au @ TiO2卵黄シェルナノ構造の調製とメチレンブルーの分解および検出へのその応用
- 炭化水素中の金ナノ粒子のサイズ制御と安定化のための分散剤としての修飾ハイパーブランチポリグリセロール
- 光熱治療および光音響イメージングのためのポリピロール被覆鉄-白金ナノ粒子の合成およびinvitro性能
- ラットにおける腹腔内および静脈内投与経路による生合成された銅および酸化亜鉛ナノ粒子の比較invivo精査
- 階層的多孔質シリカライト-1カプセル化AgNPの調製と4-ニトロフェノール還元のためのその触媒性能
- 金属および金属酸化物ナノ粒子のグリーン合成と単細胞藻類Chlamydomonasreinhardtiiに対するそれらの効果
- ナノ粒子と超音波によって制御される水の過冷却
- トランスフェリン受容体による細胞内在化が増強されたパクリタキセルベースの標的脂質ナノ粒子の抗増殖およびアポトーシス誘発能—白血病細胞での研究