


Fe3O4-Auコアシェルナノ粒子の複数の阻害剤による細胞取り込み効率の評価:結腸直腸癌細胞における特定のエンドサイトーシスを制御する可能性
要約
マグネタイト(Fe 3 O 4 )-金(Au)コアシェルナノ粒子(NP)は、独特の磁気的および光学的特性を備えています。生物学的部分と組み合わせると、これらのNPは、ドラッグデリバリーや癌の標的化などの生物医学的応用のための新しい戦略を提供することができます。ここでは、生物学的部分と組み合わせた磁気コアシェルNPシステムの制御可能な細胞取り込みのための効果的な方法を提示します。構造タンパク質であるビメンチンは、食作用に強い影響を与えることが生化学的に確認されています。さらに、ビメンチンは、生物学的部分の複数の阻害下にあるにもかかわらず、細胞への外因性物質の内在化に影響を及ぼします。この研究では、Fe 3 の細胞内在化性能を実証します O 4 -生物学的部分の組み合わせを使用して表面修飾されたAuコアシェルNP。ビメンチンタグ付きNPの光蛍光は、複数の阻害テストの下で影響を受けないままでした。これは、NPがナイスタチン、恐竜、サイトカラシンD、さらにはMuc1抗体(Ab)の影響を最小限に抑えたことを示しています。したがって、この結果は、Muc1 Abが特定の分子を標的とし、特定のエンドサイトーシスを制御できることを示しています。さらに、結腸直腸癌細胞の特定のエンドサイトーシスを制御する可能性を示しています。
はじめに
ナノマテリアルは、臨床診断と治療のための新しい道を切り開いてきました。特に、ナノ粒子(NP)は最も重要なツールの1つであり、バイオセンサー[1、2]、診断[3、4]、標的化ドラッグデリバリーシステム[5、6]などのアプリケーションで使用されています。生物医学的用途の場合、NPは通常、コア材料の表面を取り囲む有機材料で構成されます[7、8、9]。磁性材料、半導体材料、または他のタイプの材料で構成されるコア材料は、有用な物理化学的特性を持ち、外側の有機表面は、NPに化学的安定性と機能性を提供します。生物学的標的システムでの用途では、物理化学的特性だけでなく、標的化のために生体機能化された外側の有機表面も重要なパラメーターです。機能化のための標的化部分の例は、標的に特異的な抗体またはリガンドである。外表面の生体機能化された材料に応じて、NPのエンドサイトーシスメカニズムが決定されます。 NPが細胞に侵入することを可能にするメカニズムは、ナノメディシンの応用における重要性のために、多くの最近の研究の主題となっています[10、11、12、13、14、15]。
特に、磁気NPは、細胞選別[16、17]、MRI [18]、DNA分離[19]、ドラッグデリバリー[20]、温熱療法[21]、癌など、多くの特定の部位ターゲティングアプリケーションで広く使用されています。ターゲティング[22]。さまざまな磁性NPの中で、マグネタイトナノ結晶は、生体適合性と化学的安定性のために、生物医学的用途で最も広く使用されています。磁性NPを使用した生物医学的応用に多くの努力が注がれていますが、水溶液への良好な分散性、機能性、生体適合性など、いくつかの重要な問題があります。これらの問題を克服するために、多くの研究が、さまざまな官能基(たとえば、カルボキシル基やアミン基)を使用したNPの表面修飾に焦点を合わせてきました[23]。ただし、マグネタイトNPの表面への官能基の結合は、時間と手間がかかるプロセスです。この事実を考えると、Au表面は生体分子や有機材料に容易に結合できるため、コアシェルタイプのAuコーティングされた磁性NPは魅力的です。
特に、磁気コアの磁気特性-AuシェルNPは、磁気分離を可能にし、MRIイメージングの解像度を向上させ、温熱療法に適用できます。さらに、金の優れた化学結合特性は、特定の癌を標的とする受容体を介した送達システムを構築するのに有利です[24、25、26]。
過去数十年にわたって、多くの研究者が、癌を標的とする受容体を介した送達システムを報告しています[27,28,29]。
受容体を介した癌細胞の標的化は、能動的標的化の一形態です。ターゲットの選択は効果的なアクティブターゲティングの鍵であり、ターゲットは細胞外膜で過剰発現する必要があります。ほとんどの研究者は癌治療にモノクローナル抗体を使用しており、モノクローナル抗体療法を従来の化学療法と組み合わせると治療効果が大幅に向上する可能性があります[30]。モノクローナル抗体療法の成功にもかかわらず、モノクローナル抗体は癌の標的化においていくつかの制限を提示します。それらの大きなサイズ(約150 kDa)は腫瘍浸透の主要な障害であり[31、32]、それらの低い安定性と低い溶解性はそれらの広範な使用を妨げます[33]。ターゲティング担体へのそれらの付着の不均一な方向もまた、非特異的結合に対する障害と見なされます。腫瘍浸透性が改善された抗体を生成するために、さまざまな抗体フォーマットが設計およびテストされています[34]。古典的な抗体とは別に、ラクダ科の種には独特の抗体フォーマットが存在します。いわゆる重鎖抗体(HCAb)は、これらの種の末梢血および乳汁に自然に存在します。このようなHCAbの抗原結合フラグメントは、ラクダ科動物HCAb(VHH)の重鎖可変ドメイン(VH)という1つの単一ドメインで構成されています。細菌または真菌でのクローニングおよび発現後に組換えにより得られたVHHは、ナノボディと呼ばれます。分子量は11〜15 kDaで、すべてのmAbの中で最小の抗体です[35、36、37]。サイズが小さいため、孤立した場所にある抗原に対するターゲティングプローブとして適している可能性があるだけでなく、簡単に変更できる末端は、がんのターゲティングへの応用に魅力的です。
細胞の適切なターゲティングと内在化を伴うNPの効率的な送達も、送達システムの重要な要素です。ビメンチンは病原体の付着と細胞内侵入経路の構成要素として重要な役割を果たすことが報告されています。ビメンチン遺伝子発現のサイレンシングは食作用を阻害しますが[38]、切断されたビメンチンは食作用を有意に増加させるシグナルです[39]。したがって、細胞表面のビメンチンによって引き起こされる細胞食作用抵抗性を中和することは、効率的なナノ粒子送達にとって重要です。
この研究では、ナノボディタグ付きFe 3 のエンドサイトーシス経路を調査します。 O 4 -異なる長さのPEG(ポリエチレングリコール)スペーサーで修飾されたAuコアシェルNP。生化学的実験により食作用に強い影響を与えることが知られているビメンチン[39]を対照として比較し、NPの細胞内在化に効果的に作用することを確認した。また、細胞表面の糖タンパク質であり、膵臓癌、乳癌、肺癌、胃癌などのさまざまな癌で過剰発現しているMuc1は、癌を標的とするバイオマーカーとして利用されています。 Fe 3 の効率的な内部移行を確認しました O 4 AuコアシェルNPと、結腸細胞におけるMuc1受容体を介したエンドサイトーシス経路を介した癌細胞への制御可能なターゲティングの方法。
材料と方法
資料
酢酸金(III)(Au(OOCCH3)3、99.9%)はAlfaAesarから入手しました。鉄(III)アセチルアセトナート(Fe(acac) 3 を含む他の化学物質 、99.9%)、1,2-ヘキサデカンジオール(C14H29CH(OH)CH2(OH)、90%)、ポリ(エチレングリコール)-ブロック-ポリ(プロピレングリコール)-ブロック-ポリ(エチレングリコール)(PEG-PPG- PEG)、およびオクチルエーテル(C8H17OC8H17、99%)は、Sigma-Aldrichから購入し、受け取ったままの状態で使用しました。アルファ-ピリジル-2-ジスルフィド-オメガ-カルボキシスクシンイミジルエステルポリ(エチレングリコール)(OPSS-PEG-NHS)(2K、5K、および10K)はNanocsから購入しました。重曹、WST-1、クロルプロマジン、ナイスタチン、サイトカラシンD、ダイナソール、ブレフェルジンA(BFA)、モネンシン、およびトリパンブルーはSigma-Aldrichから購入しました。 Cy3とCy7.5はLumiprobeから購入しました。 Anti-Muc1 Abは、Abcam Inc.(マサチューセッツ州、ケンブリッジ)から購入しました。リン酸緩衝生理食塩水(PBS)、ダルベッコの改良イーグル培地、およびウシ胎児血清は、InvitrogenCorp。から購入しました。
Fe 3 の合成 O 4 -AuコアシェルNP
Fe 3 O 4 -AuコアシェルNPは、ナノエマルジョン法によって合成されました。コアシェルNPの合成プロセスは、次の2つのステップで構成されます。(1)Fe 3 の形成 O 4 コアNPおよび(2)磁性NP上のAuシェルのコーティング。最初のステップでは、Fe 3 O 4 NPは、Fe(acac) 3 の混合溶液から調製されました。 (0.1766gまたは0.5mmol)、1,2-ヘキサデカンジオール(0.6468gまたは2.5mmol)、およびブロック共重合体(ポリ(エチレンオキシド)-ポリ(プロピレンオキシド)-ポリ(エチレンオキシド); PEO-PPO-PEO)(オクチルエーテル中0.4〜1.2 g)。混合溶液を300℃に加熱してFe前駆体を還元した。 Fe 3 の形成 O 4 コアNPは、加熱された溶液を冷却することによって完成しました。 2番目のプロセスは、磁気コアの形成後、精製プロセスなしで継続的に実行されました。 Au前駆体(0.2338gまたは0.62mmol)および1,2-ヘキサデカンジオール(0.88 g、3.4 mmol)をFe 3 からなるエマルジョンに添加しました。 O 4 NPs、次に混合溶液を230°Cで加熱しました。室温まで冷却した後、遠心分離によってエマルジョンを沈殿させ、コアシェルNPを分離しました。
組換えAnti-Muc1-VHH5-24K10発現ベクターの構築
ポリメラーゼ連鎖反応(PCR)は、フォワードプライマー5'-CCGAATTCGCCGATGTGCAGCTGACCGAG-3 'およびリバースプライマー5'-CGG CTCGAGCTTCTTCTTCTTCTTCTTCTTCTTCTTCTTGCCTGAGGAGACGGTGACCTG-3'を使用して実施された。 PCR産物をEcoRIおよびXhoIで消化し、QIAクイックゲル抽出キット(QIAGEN、米国カリフォルニア州バレンシア)を使用してゲル精製しました。精製したPCR産物をEcoRI / XhoIで消化したpET-23a(Novagen、ダルムシュタット、ドイツ)にクローニングしました。 大腸菌 ( E.coli )DH5α(RBC Bioscience、新店、台湾)は、熱ショックによって得られたコンストラクトで形質転換され、100μg/ mLアンピシリン(Duchefa Biochemie、ハーレム、オランダ)を含むLB寒天プレート上で選択されました。
組換えタンパク質の発現と精製
組換え抗Muc1-VHH5-24 K10タンパク質を発現および精製するには、 E。コリ BL21株(RBC Bioscience、新店、台湾)をpET-23a-anti-Muc1-VHH 5-24K10で形質転換しました。次に、アンピシリン(100μg/ mL)を含むLBブロスで細菌を増殖させました。タンパク質の発現は、イソプロピルβ-d-チオガラクトシド(IPTG)(Duchefa Biochemie、Haarlem、The Netherlands)により、最終濃度0.4 mM、37°Cで5時間誘導されました。細菌ペレットを溶解緩衝液(50 mMNaH 2 )に再懸濁しました。 PO 4 、pH 8.0; 300 mMNaCl)に続いて氷上で10分間超音波処理します。超音波処理したライセートを20,000× g で遠心分離しました 4°Cで20分間、Ni-NTA His・Bindレジン(Peptron、大田、韓国)にさらしました。レジンに結合したHisタグ付きタンパク質を溶出バッファー(50 mMNaH 2 )で溶出しました。 PO 4 、pH 8.0; 300 mMNaCl; 150 mMイミダゾール)。精製したタンパク質を15%SDS-PAGEゲルで分離しました。
コアシェルFeの変更 3 O 4 -Au NP
さまざまな長さ(2、5、および10 K)のOPSS-PEG-NHSを、チオール基の活性化のために0.1M重曹に溶解しました。活性化されたOPSS-PEG-NHSが合成されたコアシェルFe 3 の溶液に添加されました O 4 -Au NPを使用し、4°Cで12時間撹拌します。活性化されたOPSS-PEG-NHSのチオール基は、コアシェルNPのAu表面に共有結合していました。次に、ナノボディ溶液(0.25 mg / mL)をPEG化Fe 3 に添加しました。 O 4 -4°Cで12時間のAuコアシェルNP。末端の10個のリジン(K)テールのアミン基は、pH8.3でOPSS-PEG-NHSのNHS基に共有結合していました。 Cy3とCy7.5は、ナノボディの残留アミン基にタグ付けされました。
内部化曲線
CT26細胞を5×10 3 で播種しました 透明な底の96ウェルプレートでウェルあたりの細胞数を250μLの培地で24時間、37°C、5%CO 2 でインキュベートします。 暗闇で。培地を除去し、50μg/ mLのCy3標識Fe 3 を含む250μLの新鮮な培地を除去しました。 O 4 -AuNPおよびPEG-Cy3またはPEG-ナノボディ-Cy3標識NPを各ウェルに添加しました。細胞をさらに異なる期間(0、10、20、30、60、120、および360分)インキュベートしました。次に、細胞をPBSで3回洗浄して遊離NPを除去し、SpectraMAX GEMINI(Molecular Devices、CA、USA)により、膜不透過性蛍光消光剤としてトリパンブルーを使用して各ウェルの蛍光を測定しました。各実験は、等量のNP(50μg/ mL)を使用して実行され、3回繰り返されました[40]。
抑制テスト
CT26細胞を5×10 3 で播種しました 透明な底の96ウェルプレートでウェルあたりの細胞数を250μLの培地で24時間、37°C、5%CO 2 でインキュベートします。 暗闇で。培地を除去し、20μg/ mLクロルプロマジン(CPZ)、50μg/ mLナイスタチン、20μg/ mLサイトカラシンD、25μg/ mLダイナソール、20μg/ mLBFA、140μg/のいずれかを含む250μLの新鮮な培養培地。 mLモネンシンまたは5μM抗Muc1Abを添加し、細胞を1時間インキュベートしました。
培地を再度除去し、50μg/ mLのCy3標識Fe 3 を含む250μLの培地を使用しました。 O 4 -Au NP、PEG-Cy3標識NP、PEG-ナノボディ-Cy3標識NP、またはビメンチン-Cy3標識NPが追加されました。
37°Cおよび5%CO 2 で1時間後 、細胞をPBSで3回洗浄して遊離NPを除去し、SpectraMAX GEMINI(Molecular Devices、CA、USA)により、膜不透過性蛍光消光剤としてトリパンブルーを使用して蛍光を測定しました。各実験は、等量のNP(50μg/ mL)を使用して実行され、4回繰り返されました。
結果と考察
コアシェルNPは、公開されている方法で合成されました[16、17]。図1a、bの透過型電子顕微鏡(TEM)観察は、Fe 3 O 4 -AuコアシェルNPは球形で、平均直径は13.5 nm、サイズ分布は狭い。
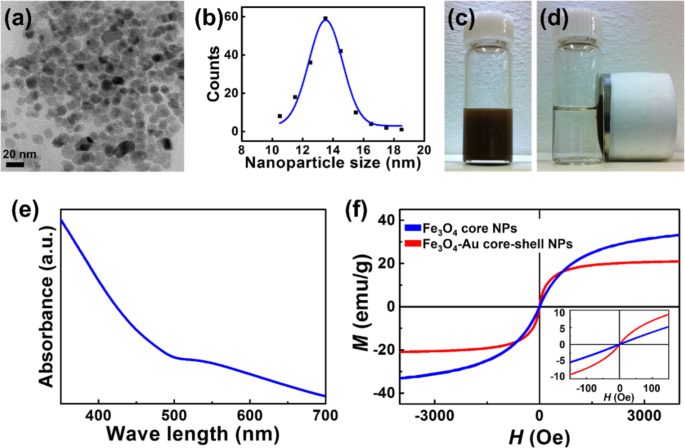
合成されたFe 3 の特性評価 O 4 -AuコアシェルNP。 a 、 b 合成されたFe 3 のTEM観察 O 4 -AuコアシェルNP。 c 、 d 外部磁場を印加する前後の水溶液中のNP。 e 合成されたコアシェルNPのUV吸光度ピークは、約530nmに現れます。 f Fe 3 の磁気ヒステリシスループ O 4 コア
コアNP(Fe 3 )の〜8.5nmからの増加 O 4 )は、コア表面に厚さ約2.5 nmのAuシェルがコーティングされているため、コアシェルNPが生成されます。 Fe 3 の高速フーリエ変換(FFT)分析による高解像度TEM画像 O 4 -AuコアシェルNPは補足情報図S1に含まれています。
有機溶媒で製造された製品は、磁気分離を使用して精製され、水中に移されました。
コアシェルNPは、NP上に存在する残留ブロック共重合体のために、表面修飾なしで水中に十分に分散し、安定していました。
図1cおよびdは、外部磁場を印加する前後の水溶液中のNPを示しています。外部磁場の下で、コアシェルNPは均一分散(図1c)から透明で透明な溶液(図1d)に急速に変化しました。
コアシェルNPの吸収帯は、UV-Vis分光法を使用して調査されました。図1eに示すように、吸光度のピークは約530 nmに現れ、NPの表面にAuが存在することを示しています(補足情報図S2には、Fe 3 のEDXデータの結果が含まれています。 O 4 -AuコアシェルNP)。サンプルが精製されたため、光学的結果はコアシェル構造の形成を示しました。
磁気ヒステリシスループは、Fe 3 の磁気特性を調査するために振動試料測定から得られました。 O 4 コアおよびコアシェルNP。両方のNPは、室温で0 Oe近くの保磁力を持つ超常磁性挙動を示しました(図1f)。
以前の研究で報告されているように、コアシェルNPの磁化率は、マグネタイトNPの磁化率よりも高かった。これは、近接効果と独特の空間構成に部分的に起因している可能性がある[41、42]。さらに、コアNPとコアシェルNPの飽和磁化は、10kOeでそれぞれ約37emu / gと約21emu / gです。 Msの違いは、コアシェルNPに非磁性成分(Au)が存在することに起因します。
VHH 5-24 K10遺伝子をインフレームでクローニングし、PCR増幅後にpET-23a-anti-Muc1-VHH 5-24 K10を生成しました(図2a)。組換えタンパク質は Eで発現された。コリ IPTGで誘導した後、pET-23a-anti-Muc1-VHH 5-24 K10で形質転換し、Ni-NTA His・BindResinで精製したBL21。組換え抗Muc1-VHH5-24K10は Eで容易に発現された。コリ 可溶性18kDaタンパク質として。 1 Lの培養物から、1±0.5mgの精製組換え抗Muc1-VHH5-24K10が得られました。
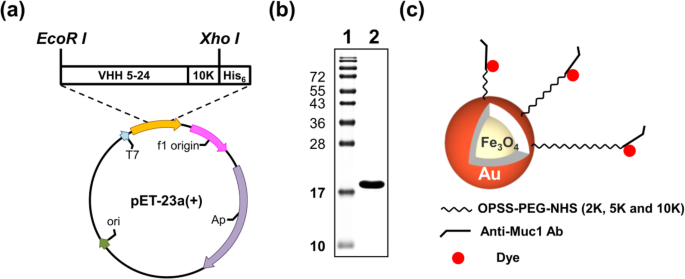
抗Muc1-VHH5-24K10融合タンパク質の発現と精製。 VHH 5-24 K10遺伝子はインフレームでクローン化され、 a を生成しました。 b 後のpET-23a-anti-Muc1-VHH5-24K10 Muc1-VHH 5-24K10融合タンパク質の精製。精製したタンパク質を15%SDS-PAGEで分離しました。レーン1タンパク質ラダー。レーン2精製タンパク質。 c この研究で使用されたPEG-ナノボディ-色素標識NPの概略図
精製したタンパク質を15%SDS-PAGEゲルで確認しました。精製されたタンパク質のクーマシーブルー染色は、それが> 95%純粋であることを明らかにしました(図2b)。合成されたFe 3 O 4 -Au NPは、PEG化、抗体タグ付け、および色素標識の3つのステップで変更されました(図2c)。各修正ステップの後、ゼータ電位を測定して、修正が成功したことを確認しました。表1は、対応するゼータ電位に対する変更の影響を示しています。裸のコアシェルNPのゼータ電位は-19.8±6.68mVでした。 NPのPEG化はOPSS-PEG-NHSを使用して実行されました。さまざまなサイズの一連のナノ複合体を生成するために、さまざまな長さ(2 K、5 K、および10 K)のOPSS-PEG-NHSが使用されました。
<図>PEG化後、ゼータ電位は低下しました(2 K、5 K、および10 Kでそれぞれ-44.9±8.19mV、-40.7±7.88 mV、および-39.6±8.74 mV)。
興味深いことに、ナノボディのタグ付け後、ゼータ電位は明らかに増加しました(2 K、5 K、および10 Kでそれぞれ-38.5±5.61mV、-23.3±8.61 mV、および-31.8±7.37 mV)。
色素タグ付け後、ゼータ電位も増加しました(2、5、および10 Kでそれぞれ-12.5±7.25mV、-17.7±3.94 mV、および-10.6±4.72 mV)。
裸のFe 3 のゼータ電位 O 4 -AuコアシェルNPは-19.8±6.68mVでした。 PEG化後、ゼータ電位は-40mV近くまで低下しました。これらの結果は、PEG分子が負に帯電したN-ヒドロキシスクシンイミド官能基を持っているため、PEG分子がコアシェルNPのAuシェルに共有結合していることを示しています。一方、ゼータ電位は、ナノボディへの色素タグ付け後に増加しました(NP-PEG2K-ナノボディでは-38.5±5.61mV、NP-PEG2K-ナノボディ-色素では-12.5±7.25mV)。組換えナノボディは末端に10個のリジンテールを持っているため、この結果は合理的です。各タイプのナノボディはゼータ電位測定によって分類され、ナノボディへの抗体の結合を決定するために、各ナノボディタイプの蛍光を測定しました。
図3に示すように、すべてのタイプのナノ粒子とナノボディが、阻害剤の制限がない場合でも、細胞への取り込みと内在化が良好であることを確認しました。細胞内在化曲線は、50μg/ mLのCy3標識Fe 3 の存在下でインキュベートした細胞から得られました。 O 4 -培地を除去し、遊離NPを洗い流し、最後にトリパンで細胞の全蛍光を測定した後の、さまざまな期間(0〜360分)のAu NP、PEG-Cy3標識NP、およびPEG-ナノボディ-Cy3標識NP青(図3)。
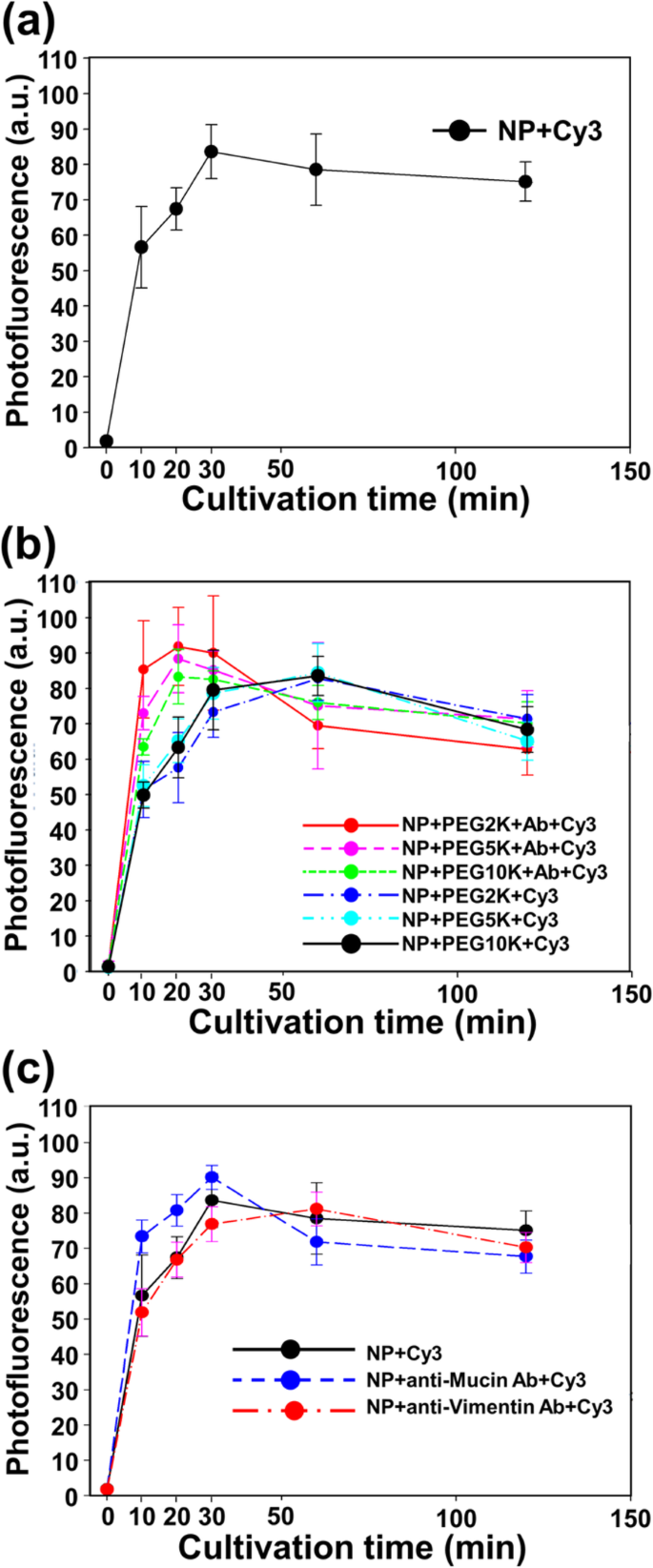
50μg/ mL a とのインキュベーション後のCT26ムチン細胞の正規化された光蛍光 Fe 3 O 4 -Au NP、 b PEG化NPとPEGナノボディタグ付きNP、および c Fe 3 O 4 -Au NP、ナノボディタグ付きNP、およびビメンチンタグ付きNP、37°C、5%CO 2 さまざまな期間(10、20、30、60、120、および360分)
蛍光強度測定の結果から、1時間以内に細胞内にNPが内在化したことがわかります(図3a)。 NPの蛍光強度は1時間以内に最大に達し、定常状態に達した後、蛍光強度は徐々に減少した。 Abの存在によってわずかな時間差がありますが、培養時間あたりの蛍光強度は、裸のNPの結果と有意な差はありませんでした(図3b)。図3cに示すように、Muc1とビメンチンの不均一なAbを使用することにより、NPの細胞取り込みと内在化においてAbの影響が無視できることが確認されました。
WST-1アッセイでは、Fe 3 の濃度と表面修飾に応じたCT26ムチン細胞の生存率(%) O 4 -AuコアシェルNPは、さまざまな曝露時間の後に推定されました(図4a、b)。 CT26細胞の生存率は、24時間および48時間の曝露後、用量を変えても、またはNPの表面修飾後でも、有意差を示さなかった。細胞生存率は、裸のFe 3 の両方で90%を超えていました O 4 -Au NP(図4a)および表面修飾NP(図4b)。
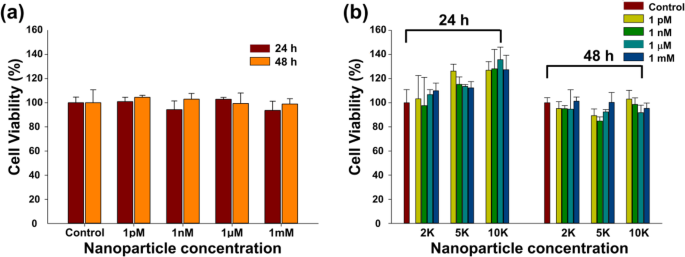
裸のFe 3 で処理されたCT26ムチン細胞の生存率 O 4 -AuコアシェルNPおよび表面修飾Fe 3 O 4 -さまざまな濃度のAuNP。 a Fe 3 O 4 -AuNPと b PEG-ナノボディ-Cy3標識NP。各実験グラフは、一連の4つの異なる実験の平均を表しています
図5は、NPがさまざまなエンドサイトーシス経路(クラスリン媒介、カベオラ媒介、および食作用/飲作用経路)を介してCT26ムチン細胞に入ったことを示しています。興味深いことに、図5bは、抗Muc1Abが主にPEG-ナノボディ-Cy3標識NPのエンドサイトーシスにも影響を及ぼしたことを示しています。 NPの内在化の経路を理解するために、特定の化学阻害剤でエンドサイトーシス経路を阻害しようとしました(図5)。エンドサイトーシスの経路は、クラスリン介在性、カベオラ介在性、およびマクロ飲作用/食作用の3つのタイプに分類されることがよく知られています。
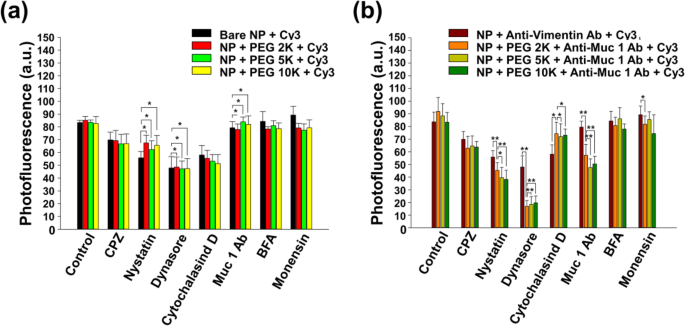
化学エンドサイトーシス阻害剤で1時間処理し、50μg/ mLでインキュベートしたCT26ムチン細胞からの正規化された光蛍光 a ベアコアシェルNPおよび b 5%CO 2 中の37°Cでのナノボディタグ付きNP 30分。内在化に統計的影響を与える阻害剤(学生の t テスト、 p (*)<0.05および p (**)<0.01)は黒いアスタリスクでマークされています
この研究では、阻害剤は、ナノボディタグ付きNPの内部移行を調査するための最初のアプローチとして使用されました。 CPZ(クラスリン媒介エンドサイトーシス阻害剤)、ナイスタチン(カベオラ媒介エンドサイトーシス阻害剤)、ダイナソール(ダイナミン阻害剤)、サイトカラシンD(食作用/マクロピノサイトーシス阻害剤)、BFA(ゴルジ装置破壊剤)、モネンシン(リソソーム阻害剤)、または抗Muc1 Ab(受容体/トランスポーター特異的競合物質)を細胞と1時間インキュベートしました。 CPZ、ナイスタチン、恐竜、サイトカラシンDは、NPのエンドサイトーシスに影響を及ぼしました(図5a)。
標的化部分は、癌治療において非常に重要である癌標的化の成功への鍵です。ターゲティングの場合、効果的な表面修飾は、治療効率を高め、副作用を制限するために非常に重要です。ナノボディタグ付きFe 3 O 4 -AuコアシェルNPは、合成されたNPと組換えナノボディから正常に作成されました。表1は、各変更ステップが正常に実行されたことを明確に示しています。
細胞の生存率は、ナノマテリアルの生物学的応用に不可欠な要素の1つです。細胞生存率は、ベアコアシェルNPおよび修飾NPで90%を超えていました(図4a、b)。これらの結果は、裸のFe 3 O 4 -Au NPおよび修飾NPは、濃度および修飾に依存する有意な細胞毒性を引き起こさず、修飾NPは生物学的用途に適していました。
NPの内在化効率と阻害剤効果の研究は、NPが細胞に入るメカニズムを理解するための重要な情報を提供します。 PEG化されたNPは、裸のNPと比較して細胞への内在化が比較的遅かったが、ナノボディタグ付きNPは、裸のNPよりもわずかに速く細胞への内在化された(図3a、b)。 PEG化はNPの内部移行を防ぐためのよく知られた表面修飾方法であるため、PEG化されたNPの内部移行傾向は簡単に説明できます。さらに、ナノボディはNPのエンドサイトーシスを誘発しました。ナノボディの特異性を確認するために、ビメンチンAbタグ付きNPの内在化率を確認しました。興味深いことに、ビメンチンAbタグ付きNPは細胞内在化を促進しませんでした(図3c)。
これらの結果は、ナノボディがナノ材料のCT26ムチン細胞への内在化を効果的に誘導できることを示しており、内在化傾向はNPの外膜の特定の修飾によって制御できることを意味します。さらに、ナノボディタグ付きNPのエンドサイトーシスのメカニズムは、阻害試験と共焦点顕微鏡イメージングによって明確に示されました。ナノボディタグ付きNPとタグなしNPの両方の光蛍光は、CPZ、ナイスタチン、または恐竜と一緒に培養した場合、同様の減少値を示しました(図5a、b)。 CPZ、ナイスタチン、およびダイナソーアは、それぞれ、クラスリン媒介エンドサイトーシス、カベオラ媒介エンドサイトーシス、および親膜からの新生小胞の出芽および切断に関与する大きなGTPアーゼであるダイナミンを阻害する役割を果たします。これにより、ダイナミンはクラスリン介在性およびカベオラ介在性エンドサイトーシスの小胞の生成と密接に関連しているため、両方の場合の光蛍光値は急速に減少しました。図5aに示すように、タグなし(図5a)とナノボディタグ付きNP(図5b)の両方で、光蛍光の急激な減少が見られました。
特に、ナノボディタグ付きNPは、CPZ、ナイスタチン、および恐竜で有意に低い光蛍光値を示しました。さらに、食作用のためにアクチンフィラメントの重合をブロックする細胞透過性毒素であるサイトカラシンDに適用した場合、タグなしNPはナノボディタグ付きNPよりも強く影響を受けることを確認しました[43]。これらの結果は、タグ付けされていないNPが、クラスリンを介したエンドサイトーシス、カベオラを介したエンドサイトーシス、食作用などの複数のメカニズムによって内在化されたことを意味します。その結果、ナノボディタグ付きNPの細胞内在化は、クラスリン媒介およびカベオラ媒介エンドサイトーシスに依存します。さらに、細胞をMuc1抗体で培養すると、ナノボディタグ付きNPの細胞取り込み量が大幅に減少しました(図5b)。この結果は、遊離のMuc1抗体が、CT26細胞膜への付着において、修飾されたNP上のナノボディの競合物質としての役割を果たし、Muc1抗体が修飾されたNPの細胞内在化において重要な役割を果たすことを示しています。特異なことに、ビメンチンAbタグ付きNPは、阻害能力の点でナノボディタグ付きNPと比較して明らかな違いを示しました。ビメンチンタグ付きNPの光蛍光は、複数の阻害テストの下で影響を受けないままでした。これは、NPがナイスタチン、恐竜、サイトカラシンD、さらにはMuc1Abによる影響を最小限に抑えたことを示しています。この現象は、食作用に強力な影響を与えることが生化学的に確認されているビメンチンの有効性の証拠である可能性があります[39]。したがって、この結果は、Muc1 Abが特定の分子を標的とし、特定のエンドサイトーシスを制御できることを示しています。
図6に示すように、細胞をCy7.5標識ベアコアシェルNPおよびPEGナノボディタグ付きNPで処理した場合にも同様の結果が得られ、恐竜阻害がない場合の両方のケースで同様の細胞取り込みが示されました。恐竜の抑制は、裸のNPと比較して、PEG-ナノボディタグ付きNPの細胞内在化を明らかに低下させました(図6b、下の行)。これらの結果は、2つのエンドサイトーシスメカニズムがあることを意味します。それは、裸のNPの非特異的エンドサイトーシスと、ダイナミン分子を介したナノボディタグ付きNPの制限されたエンドサイトーシスです。ナノボディが外部の細胞膜に付着すると、クラスリンとカベオラを介したメカニズムが同時に活性化されるため、ナノボディのタグが付けられたNPは細胞膜を簡単に通過できます。したがって、主なメカニズムは、CT26ムチン細胞におけるナノボディタグ付きNPの内在化のためのクラスリンおよびカベオラを介したエンドサイトーシスの両方であると推測できます。
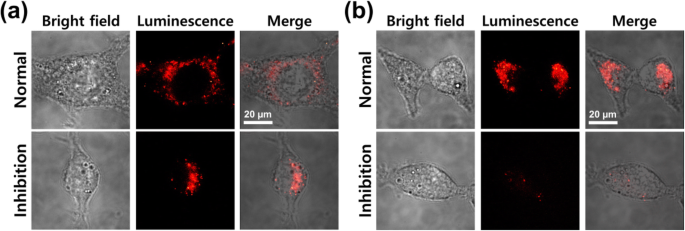
a とインキュベートしたCT26ムチン細胞の共焦点顕微鏡イメージング Fe 3 O 4 -AuNPと b PEG-ナノボディタグ付きNP、37°C、5%CO 2 で1時間 インキュベーター、恐竜抑制の前後(1 ng / mL)
結論
癌を標的とし、薬物、遺伝子、ペプチドなどの外因性物質を制御可能に挿入するためのナノ材料は、生物医学的応用における重要な進歩です。これらの馴染みのある、しかし創造的な概念は、新しい治療法の戦略を提供することができます。この論文では、Fe 3 の細胞取り込みの強化を実証しました O 4 -Muc1抗体でPEG化した後のAuコアシェルNP。ナノボディタグ付きNPの主なエンドサイトーシスメカニズムが実証され、結腸直腸癌細胞における制御可能な特定のエンドサイトーシスの可能性が示されました。これらの調査結果は、ナノボディタグ付きNPと結腸直腸癌細胞の間のターゲティングに関する洞察を提供し、高効率のターゲティングキャリアの設計を支援します。
データと資料の可用性
この調査中に生成または分析されたすべてのデータは、この公開された記事に含まれています。
略語
- Fe 3 O 4 :
-
マグネタイト
- Au:
-
ゴールド
- NP:
-
ナノ粒子
- Ab:
-
抗体
- HCAbs:
-
重鎖抗体
- VHH:
-
ラクダHCAbのVH
- PEG:
-
ポリ(エチレングリコール)
- PPG:
-
ポリ(ポリプロピレングリコール)
- PCR:
-
ポリメラーゼ連鎖反応
- TEM:
-
透過型電子顕微鏡
- PEO-PPO-PEO:
-
ポリ(エチレンオキシド)-ポリ(プロピレンオキシド)-ポリ(エチレンオキシド)
- OPSS-PEG-NHS:
-
オルトピリジルジスルフィドPDPPEGスクシンイミジルエステル
- BFA:
-
ブレフェルディンA
- CPZ:
-
クロルプロマジン
ナノマテリアル
- 高効率グラフェン太陽電池
- 癌治療のためのナノ粒子:現在の進歩と課題
- 癌用途のための細胞ベースのドラッグデリバリー
- ナノテクニックは癌幹細胞を不活性化する
- 多機能GaN / Feナノ粒子による内皮細胞の標的化
- 炭化水素中の金ナノ粒子のサイズ制御と安定化のための分散剤としての修飾ハイパーブランチポリグリセロール
- ペロブスカイト太陽電池の変換効率に及ぼすTiO2コンパクト層に埋め込まれた異なるサイズと濃度のAgナノ粒子の影響
- トランスフェリン受容体による細胞内在化が増強されたパクリタキセルベースの標的脂質ナノ粒子の抗増殖およびアポトーシス誘発能—白血病細胞での研究
- 形状選択的な細胞取り込みのための成長する金ナノ構造
- リトコール酸で修飾された金ナノ粒子の肝臓癌細胞に対するアポトーシス効果
- 骨芽細胞様3T3-E1細胞におけるジルコニアナノ粒子誘発毒性効果