


阻害されたMicroRNA-301は、PTENを上昇させることにより、食道扁平上皮癌の血管新生と細胞増殖を抑制します
要約
目的
食道扁平上皮癌(ESCC)は、早期転移と診断の遅れが特徴です。 MicroRNA-301(miR-301)は、さまざまな癌に関与することが知られています。それにもかかわらず、ESCCに対するmiR-301の影響は未解明のままです。したがって、ESCCの進行におけるmiR-301の役割を調査することを目指しています。
メソッド
ESCC組織および細胞株におけるmiR-301およびホスファターゼおよびテンシンホモログ(PTEN)の発現を評価しました。次に、スクリーニングした細胞を変更したmiR-301またはPTENオリゴヌクレオチドとプラスミドで処理し、ESCC細胞のコロニー形成能、細胞生存率、遊走、浸潤、細胞周期分布、アポトーシスを評価しました。さらに、腫瘍の成長と微小血管密度(MVD)も評価され、miR-301とPTENのターゲティング関係が確認されました。
結果
MiR-301はアップレギュレーションされ、PTENはESCC組織および細胞でダウンレギュレーションされました。 KYSE30細胞とEca109細胞が機能アッセイのために選択されました。 KYSE30細胞では、miR-301の阻害または過剰発現したPTENが細胞の悪性行動を抑制し、PTENの沈黙がESCCの進行に対するmiR-301阻害の影響を排除しました。 Eca109細胞では、miR-301の過剰発現またはPTEN阻害が細胞の悪性行動を促進し、PTENの過剰発現はESCCの進行に対するmiR-301の上昇の影響を逆転させました。インビボアッセイは、miR-301阻害またはPTEN過剰発現がESCC腫瘍増殖およびMVDを抑制し、miR-301上昇またはPTEN減少が逆の効果を有することを明らかにした。さらに、PTENはmiR-301の標的になりました。
結論
まとめると、私たちの研究の結果は、miR-301がESCCでのPTEN発現の調節を介して細胞増殖、転移、血管新生に影響を与えることを明らかにしました。
はじめに
世界で8番目に多い癌である食道癌(EC)は、高い死亡率と予後不良を伴う重大な悪性腫瘍です[1]。全EC症例の約90%を占める食道扁平上皮癌(ESCC)は、中国におけるECの主な形態です[2]。低い社会経済的地位、喫煙、アルコール消費、栄養失調、ニトロソアミンが豊富な、またはマイコトキシンで汚染された食品を含む複数の原因がESCCの発生につながります[3]。臨床転帰と投与が促進されているにもかかわらず、ESCC患者の予後は依然として不良であり、5年生存率は15〜25%です[4]。したがって、ESCC患者にさらに効果的な治療法を提供するために、ESCCの開発においてバイオマーカーとして機能する可能性のある癌遺伝子または腫瘍抑制遺伝子を確認することが重要です。
マイクロRNA(miRNA)は、遺伝子発現の調節に不可欠な役割を果たす小さな非コードRNAであり[5]、mRNAの安定性とmRNAの能力を調節することで腫瘍の進行に影響を与える能力があることが明らかになっています[6]。 miR-4324 [7]、miR-889-3p [8]、miR-9 [9]などのmiRNAの量は、ESCCのプロセスに関連していることがわかっています。 MiR-301は、ヒトゲノムの17q22-23に位置するfam33a転写ユニットによって形成されるmiRNAのメンバーです。 miR-301の過剰発現は以前に同定されており、これはヒトの疾患に関係していることを反映しています[6、10]。ただし、miR-301の機能メカニズムはESCCで明らかにされていません。さらに、ホスファターゼおよびテンシンホモログ(PTEN)は、腫瘍で頻繁に破壊され、腫瘍の抑制的役割を果たす癌患者の生殖細胞変異の標的となることが確認されています[11]。 PTENの調節不全はESCCの発症と相関していることが確認されています[12]。興味深いことに、最近の研究により、miR-301は非小細胞肺癌のPTENを標的としていることが明らかになりました[13]。ただし、ESCC開発におけるmiR-301とPTENのこのターゲティング関係は、まだ明らかにされていません。私たちの研究は、ESCCの進行に対するmiR-301とPTENの影響に焦点を当てました。これらはほとんど不明であり、目新しいものです。 miR-301は、PTENの発現を調節することにより、ESCCの血管新生と細胞増殖に影響を与える可能性があると推測しました。
材料と方法
倫理声明
書面による情報に基づく同意は、研究の前にすべての患者から取得されました。この研究のプロトコルは、吉林大学第二病院の倫理委員会によって承認され、ヘルシンキ宣言の人間を対象とした医学研究の倫理原則に基づいています。動物実験は、国立衛生研究所が発行した実験動物の管理と使用に関するガイドと厳密に一致していました。動物実験のプロトコルは、吉林大学第2病院の施設内動物管理使用委員会によって承認されました。
調査対象
1月から吉林大学第2病院の胸部外科で食道切除術を受けたESCC患者(男性78人と女性32人)から、ESCC組織と隣接する正常組織(腫瘍から> 5cm)のサンプル110個を収集しました。 2015年から2017年12月。110人の患者のうち、84例> 60年、26例≤60年でした。腫瘍サイズ:65例≥5cmおよび45例<5cm;リンパ節転移(LNM)のない71例とLNMのある39例。腫瘍、リンパ節転移(TNM)期:60例がI + II期、50例がIII期でした。腫瘍の位置:13例が上部ESCCで、97例が中下部ESCCでした。患者は全員ESCCと診断され、以前は放射線療法や化学療法を受け入れていませんでした。腫瘍は完全に切除され、病理学によって負の外科的切除断端が確認された。 2009年に国際対がん連合(UICC)によって提案されたESCCの病期分類基準[14]によれば、患者の術後の病理学的病期分類はpT1-4N1-2(I-IIIb)病期として特定されました。手術後の患者に重大な合併症はなく、周術期死亡は除外されました。
逆転写定量的ポリメラーゼ連鎖反応(RT-qPCR)
Trizolキット(Invitrogen Inc.、Carlsbad、CA、USA)を使用して、組織および細胞のトータルRNAを抽出しました。 RNAの濃度と品質を測定しました。 RNAプライマー(表1)は、TaKaRa Biotechnology Co.、Ltd。(Liaoning China)によって設計および合成されました。タカラPrimeScript™RT試薬キットとgDNA Eraser(Takara)の指示に基づいて、RNAをcDNAに逆転写しました。 Power PCR SYBRグリーンPCRマスターミックス(Takara)を使用してLight Cycler 480II(Roche)でqPCRを実行しました。 miR-301のローディングコントロールとしてU6を使用し、PTENの内部参照としてβ-アクチンを使用しました。データは2 -△△Ct を使用して分析されました メソッド[15]。
ウエスタンブロット分析
RIPA溶解バッファー(Beyotime Institute of Biotechnology、上海、中国)を使用して細胞および組織中の総タンパク質を抽出し、BCAタンパク質アッセイキット(Beyotime)によってタンパク質を定量しました。各サンプルのタンパク質濃度を測定し、10%ドデシル硫酸ナトリウム-ポリアクリルアミドゲル電気泳動を行いました。サンプルをニトロセルロースメンブレンに転写し、5%スキムミルクパウダーで4°Cで一晩ブロックしました。その後、メンブレンに一次抗体PTENおよびβ-アクチン(両方とも1:500およびSanta Cruz Biotechnology Inc、CA、USAから)を一晩インキュベートするために補充し、次にそれぞれの二次抗体を添加して1時間インキュベートしました。強化された化学発光試薬(Pierce Chemical Inc.、Dallas、TX、USA)に1分間浸した後、メンブレンを暗い環境にさらし、LAS4000ミニ化学発光イメージャーを使用して現像しました。グレー値は、コントロールとしてβ-アクチンを使用したイメージングシステムソフトウェアによって評価されました。したがって、発現された最終的な相対タンパク質が得られた。タンパク質バンドはImageJ2xソフトウェアによって分析されました。
デュアルルシフェラーゼレポーター遺伝子アッセイ
PTENの3 '非翻訳領域(UTR)配列は、miR-301と相互作用すると予測されました。または、予測された標的部位内の変異配列が合成され、pGL3コントロールルシフェラーゼレポーターベクター(Promega、 WI、USA)。次に、pGL3-PTEN-wtおよびpGL3-PTEN-mutベクターを作成しました。 miR-301ミミックおよびミミックNCを含む正しく同定されたwtおよびmutルシフェラーゼレポータープラスミドを、KYSE30およびEca109細胞に48時間コトランスフェクトしました。続いて、細胞を溶解し、ルシフェラーゼ活性をそれぞれルシフェラーゼ検出キット(Promega)で測定しました。
細胞培養、グループ化、トランスフェクション
ESCC細胞株(KYSE-150、KYSE-30、Eca109およびKYSE-70)は、上海生化学細胞生物学研究所、中国科学院(上海、中国)から入手し、ヒト食道上皮細胞(HEEC)を入手しました。 Mingzhou Biotechnology Co.、Ltd。(Zhejiang、China)から。細胞は、10%ウシ胎児血清(FBS、Life Technologies、USA)、100 units / mlペニシリンGナトリウム(Sigma)および100μg/ mlストレプトマイシン硫酸塩(Sigma)を添加したRPMI 1640培地(Invitrogen)で培養しました。各細胞株におけるMiR-301発現とPTENのmRNA発現をRT-qPCRで測定し、相対発現が最も高い細胞株と最も低い細胞株をその後の細胞実験に選択しました。
KYSE-30細胞を7つのグループに分け、それぞれ、miR-301阻害剤、阻害剤陰性対照(NC)、pcDNA-PTEN(過剰発現(oe)-PTENと名付けられた)、pcDNA-NC(oe-NCと名付けられた)、 miR-301阻害剤+低分子干渉RNA(si)-PTENまたはmiR-301阻害剤+ si-NC。 Eca109細胞も7つのグループに分けられ、miR-301 mimic、mimic NC、si-PTEN、si-NC、miR-301 mimic + oe-PTEN、miR-301 mimic + oe-NCで個別に処理されました。阻害剤NC、miR-301阻害剤、miR-301ミミック、ミミックNC、si-NC、およびsi-PTENは、GenePharma Ltd.、Company(Shanghai、China)から購入しました。 pcDNA-PTEN NCおよびpcDNA-PTENは、(Shanghai Sangon Bio-technology Corporation(Shanghai、China))から入手しました。細胞のコンフルエンスが60%に達したときに、リポフェクタミン2000(Invitrogen)によって細胞をESCC細胞に一過性にトランスフェクトしました。
細胞カウントキット(CCK-8)アッセイ
細胞を96ウェルプレートに播種しました(1×10 3 細胞/ウェル)および異なる期間インキュベートした。 24時間、48時間、72時間、および96時間インキュベートした後、各ウェルに10μLのCCK-8溶液(5 mg / mL)を補充し、次に各グループの細胞を光なしで37°Cでインキュベートしました。 2時間の暴露。 450 nmでの光学密度(OD)値は、マイクロプレートリーダー(Bio-Rad Laboratories、Hercules、CA、USA)によって分析されました。
コロニー形成アッセイ
トランスフェクション後、細胞を6ウェルプレートに500細胞/ウェルで播種し、14日間培養しました。コロニーをメタノールで固定し、0.5%クリスタルバイオレットを使用して染色し、倒立顕微鏡でカウントしました。
トランスウェルアッセイ
セル(5×10 3 )RPMI 1640培地で培養したものを、コーティングされていない、またはマトリゲルでコーティングされたメンブレン(Corning、NY、USA)を備えたTranswellデバイスの頂端チャンバーに播種しました。 24時間後、頂端チャンバー上の細胞を除去し、下側に残っている細胞を固定し、0.1%クリスタルバイオレットを使用して染色しました。顕微鏡(オリンパス株式会社、東京、日本)を使用して、3つの確率場を数え、細胞数を計算しました。
フローサイトメトリー
細胞周期とアポトーシスはフローサイトメトリーによって評価されました。アネキシンV-フルオレセインイソチオシアネート(10 µL)およびヨウ化プロピジウム(PI; 5 µL、Sigma)を細胞とインキュベートしました(5×10 5 細胞/ウェル)4°Cで30分間暗所で。アポトーシス細胞のパーセンテージは、FlowJoバージョン10ソフトウェア(FlowJo LLC、OR、USA)を備えたフローサイトメーター(BD Biosciences、CA、USA)を使用して計算されました。
細胞周期を評価するために、細胞(5×10 5 細胞/ウェル)を75%エタノールで4°Cで一晩固定し、5 µlのPI /リボヌクレアーゼA(Sigma)で4°Cで30分間暗所で染色しました。データはフローサイトメーター(BDBiosciences)で分析されました。各サンプルの蛍光シグナル(14,000)は、ModFit LTバージョン3.2ソフトウェア(Verity Software House、Inc.、ME、USA)を使用して収集および計算されました。
ヌードマウスの皮下腫瘍形成
42匹の雌のBALB / c-nuヌードマウス(4 w、体重16〜24 g)は、実験動物センター吉林大学(中国、長春)から入手しました。ヌードマウスは14のグループに分けられました( n =3)。 7つのグループのヌードマウスは、細胞のグループ化に従って、それぞれ、KYSE-30細胞を注射され、残りの7つのグループのヌードマウスは、グループ化に基づいて、Eca109細胞を別々に注射された。トランスフェクトされたKYSE-30およびEca109細胞の濃度は5×10 6 に調整されました 細胞/100μL。ヌードマウスを固定し、滅菌条件下で対応するESCC細胞を皮下注射した。腫瘍の最大の長さ(L)と幅(W)を毎週測定し、腫瘍の体積( V )=1/2× L × W 2 。ヌードマウスは5回目で安楽死させた 腫瘍を切除した注射の週、および腫瘍の重さを量り、写真を撮った。腫瘍形成率は、皮下腫瘍を有するマウスの数/グループ×100%に注射されたヌードマウスの総数として計算された。注射時間を横軸とし、腫瘍サイズを縦軸とした。したがって、腫瘍増殖曲線がグラフ化されました。
免疫組織化学的染色
ヌードマウスの腫瘍組織を10%ホルムアルデヒドで固定し、パラフィンで包埋し、4μmに切断しました。次に、切片を60°Cで2時間乾燥し、キシレンで脱ロウし、勾配エタノールで脱水し、50μLの3%H 2 とインキュベートしました。 O 2 10分間。その後、切片を0.01 Mクエン酸緩衝液に浸し、95°Cで20分間煮沸し、37°Cで10分間通常のヤギ血清作業溶液でブロックし、CD 34 を添加しました。 (1:100、Santa Cruz)4°Cで一晩。その後、切片にHRP標識ヤギ抗ウサギ/マウスIgGポリマー(ZSGB-Bio、北京、中国)を補充し、ヘマトキシリンで対比染色し、脱水および透過処理した後、中性バルサムで密封しました。 NCとして一次抗体の代わりにPBSを使用しました。微小血管密度(MVD)の測定:切片を低倍率の顕微鏡で観察した。内皮細胞または内皮細胞クラスターが茶色がかった黄色に染色され、周囲の腫瘍細胞と有意に区別され、結合組織が微小血管として採取された。分岐構造は、切断された場合も血管と見なされ、内腔サイズ> 8の赤血球、筋層、またはより厚い内腔の血管は除外されました。 3つの高視野の微小血管の数を記録し、平均数は各症例のMVDでした。
統計分析
すべての統計分析は、SPSSバージョンSPSS 21.0ソフトウェア(IBM Corp. Armonk、NY、USA)を使用して実行され、Graphpad Prism Software6.0で提供されました。データは平均±標準偏差として表されました。 2つの独立したグループ間の差異は、スチューデントのt検定でテストされました。 3つ以上のグループを比較するために一元配置分散分析を実行しました。 P 値<0.05は統計的に有意な差を示しました。
結果
MiR-301は高度に発現していますが、PTENはESCC組織および細胞では発現が不十分です
ESCC組織および隣接する正常組織におけるMiR-301およびPTENの発現を、RT-qPCRおよびウエスタンブロット分析を使用して評価し、ESCCにおけるそれらの役割を明らかにしました。(図1a–c)miR-301はアップレギュレーションされ、PTENはアップレギュレーションされました。 ESCC組織でダウンレギュレーションされました。 miR-301またはPTEN発現の中央値に従って患者を低発現群と高発現群に分け、miR-301またはPTEN発現とESCC患者の臨床病理学的特徴との相関関係を分析しました。結果は、miR-301 / PTENの発現は年齢、性別、腫瘍の大きさ、位置、分化とは関係がなく、ESCC患者のTNM病期とLNMとは相関していることを示しました(表2)。
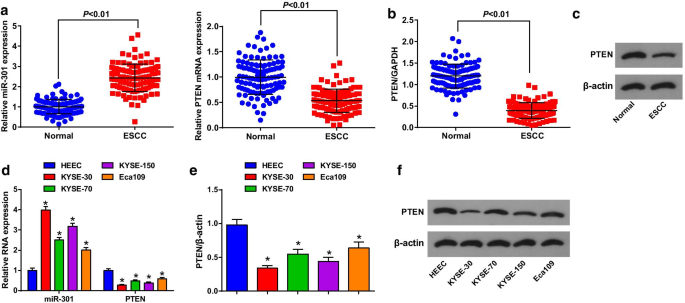
MiR-301は高度に発現していますが、PTENはESCC組織および細胞ではほとんど発現していません。 a RT-qPCRを使用して検出されたESCC組織におけるmiR-301の発現とPTENのmRNA発現。 b ウエスタンブロット分析を使用して検出されたESCC組織におけるPTENのタンパク質発現。 c ウエスタンブロット分析におけるESCC組織のPTENのタンパク質バンド。 d RT-qPCRを使用して検出されたESCC細胞株におけるmiR-301の発現とPTENのmRNA発現。 e ウエスタンブロット分析を使用して検出されたESCC細胞株におけるPTENのタンパク質発現。 f ウエスタンブロット分析におけるPTENのタンパク質バンド。 * P <0.05対HEEC。データは平均±標準偏差として表され、2つのグループ間の比較のためにt検定が実行されました
次に、4つのESCC細胞株およびHEECにおけるmiR-301およびPTENの発現を、RT-qPCRおよびウエスタンブロット分析を使用して決定しました。 ESCC細胞株ではmiR-301がアップレギュレーションされ、PTENがダウンレギュレーションされ、その中でKYSE-30が最も高いmiR-301発現と最も低いPTEN発現を示し、Eca109は逆の傾向を示しました。したがって、細胞実験では、KYSE-30細胞株をダウンレギュレーションされたmiR-301 /過剰発現PTENで処理し、Eca109細胞株を過剰発現したmiR-301 /サイレンシングPTENで処理しました。
PTENはmiR-301のターゲットになっています
バイオインフォマティクスソフトウェア(http://www.microrna.org/)は、PTENがmiR-301の標的遺伝子であると予測しました(図2a)。さらに、デュアルルシフェラーゼレポーター遺伝子アッセイにより、PTEN-wtベクターとmiR-301ミミックをコトランスフェクトしたESCC細胞では、PTEN-mutベクターとmiR-301ミミックをコトランスフェクトした細胞と比較して、ルシフェラーゼ活性が有意に低下したことが確認されました。 miR-301が特にPTENに結合する可能性があることを意味します(図2b、c)。
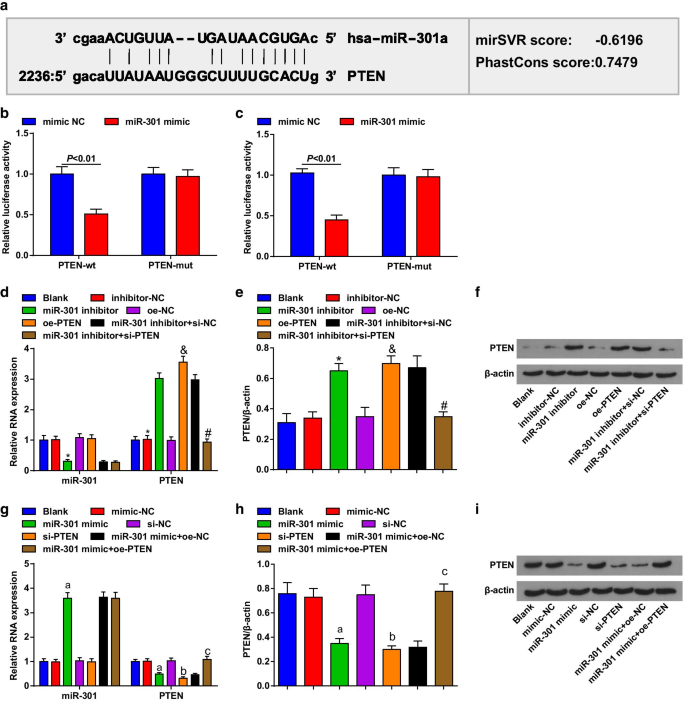
PTENはmiR-301の標的遺伝子です。 a miR-301とPTENの結合部位は、オンライン予測ソフトウェアによって予測されました。 b KYSE-30細胞におけるmiR-301とPTENの標的関係は、デュアルルシフェラーゼレポーター遺伝子アッセイによって評価されました。 c Eca109細胞におけるmiR-301とPTENの標的関係は、デュアルルシフェラーゼレポーター遺伝子アッセイによって評価されました。 d miR-310のダウンレギュレーションまたはPTENのアップレギュレーション後にRT-qPCRを使用して検出されたKYSE-30細胞におけるmiR-301の発現およびPTENmRNAの発現。 e miR-310のダウンレギュレーションまたはPTENのアップレギュレーション後にウエスタンブロット分析を使用して検出されたKYSE-30細胞におけるPTENのタンパク質発現。 f miR-310のダウンレギュレーションまたはPTENのアップレギュレーション後のウエスタンブロット分析におけるKYSE-30細胞のPTENのタンパク質バンド。 g miR-310のアップレギュレーションまたはPTENのダウンレギュレーション後にRT-qPCRを使用して検出されたEca109細胞におけるmiR-301の発現およびPTENのmRNA発現。 h miR-310のアップレギュレーションまたはPTENのダウンレギュレーション後にウエスタンブロット分析を使用して検出されたEca109細胞におけるPTENのタンパク質発現。 i miR-310のアップレギュレーションまたはPTENのダウンレギュレーション後のウエスタンブロット分析におけるEca109細胞のPTENのタンパク質バンド。 * P <0.05対阻害剤-NCグループ、および P <0.05対oe-NCグループ、 # P <0.05対miR-301阻害剤+ si-NCグループ、 P <0.05対mimic-NCグループ、b P <0.05対si-NCグループ、c P <0.05対miR-301ミミック+ oe-NCグループ。 N =3.データは平均±標準偏差として表され、2つのグループ間の比較のためにt検定が実行されました。複数のグループ間の比較にANOVAを使用しました
RT-qPCRおよびウエスタンブロット分析を使用して、トランスフェクトされた細胞におけるmiR-301およびPTENの発現を評価し、KYSE-30細胞(図2d–f)では、miR-301阻害剤で処理された細胞がmiR-301をダウンレギュレーションすることがわかりました。 、PTENをアップレギュレーションしている間; pcDNA-PTEN(oe-PTEN)で処理された細胞はPTEN発現を上昇させ、si-PTENはPTEN発現に対するmiR-301阻害剤の効果を逆転させました。 Eca109細胞(図2g–i)では、miR-301で処理された細胞はアップレギュレーションされたmiR-301を模倣しますが、ダウンレギュレーションされたPTENを模倣します。 si-PTENで処理された細胞はPTEN発現を減少させ、pcDNA-PTEN(oe-PTEN)はPTEN発現におけるmiR-301模倣物の阻害的役割を逆転させました。これらのデータは、miR-301がPTENを標的にしていることを示唆しています。
抑制されたmiR-301または過剰発現されたPTENは、ESCC細胞の生存を制限します。 miR-301の上昇またはPTENの低下はESCC細胞の生存率を促進します
ESCC細胞の細胞生存率は、コロニー形成とCCK-8アッセイを使用して評価されました。その結果、KYSE-30細胞株(図3a–c)では、miR-301阻害剤またはoe-PTENのトランスフェクションにより、コロニー形成能と細胞生存率が抑制されたことが明らかになりました。サイレンシングされたPTENのトランスフェクションにより、ESCC細胞の生存率に対するmiR-301阻害剤の影響が排除されました。 Eca109細胞株(図3d–f)では、miR-301ミミックまたはsi-PTENのトランスフェクションにより、コロニー形成能と細胞生存率が促進されました。 PTENの過剰発現は、Eca109細胞のコロニー形成能力と生存率におけるmiR-301上昇の促進的役割を逆転させました。これらの結果は、miR-301ノックダウンまたはPTENの過剰発現が、miR-301の上昇またはPTEN阻害によって促進されたESCC細胞の生存率を抑制したことを示唆しています。
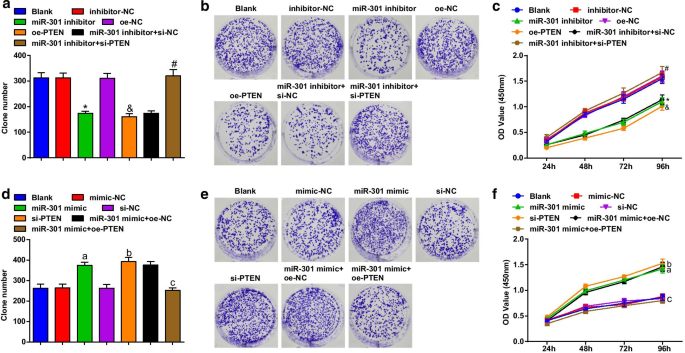
阻害されたmiR-301または過剰発現されたPTENは、ESCC細胞の生存を制限します。 miR-301の上昇またはPTENの低下は、ESCC細胞の生存率を促進します。 a miR-310のダウンレギュレーションまたはPTENのアップレギュレーション後にコロニー形成アッセイを使用して検出されたトランスフェクション後のKYSE-30細胞のコロニー数。 b miR-310ダウンレギュレーションまたはPTENアップレギュレーション後のコロニー形成アッセイを使用して検出されたトランスフェクション後のKYSE-30細胞のコロニー形成能力。 c miR-310のダウンレギュレーションまたはPTENのアップレギュレーション後にCCK-8アッセイを使用して検出されたトランスフェクション後のKYSE-30細胞の生存率。 d miR-310のアップレギュレーションまたはPTENのダウンレギュレーション後にコロニー形成アッセイを使用して検出されたトランスフェクション後のEca109細胞のコロニー数。 e miR-310アップレギュレーションまたはPTENダウンレギュレーション後のコロニー形成アッセイを使用して検出されたトランスフェクション後のEca109細胞のコロニー形成能力。 f miR-310のアップレギュレーションまたはPTENのダウンレギュレーション後にCCK-8アッセイを使用して検出されたトランスフェクション後のEca109細胞の生存率。 * P <0.05対阻害剤-NCグループ; & P <0.05対oe-NCグループ; # P <0.05対miR-301阻害剤+ si-NCグループ; P <0.05対mimic-NCグループ。 b P <0.05対si-NCグループ; c P <0.05対miR-301ミミック+ oe-NCグループ、 N =3.データは平均±標準偏差として表され、ANOVAは複数のグループ間の比較に使用されました
抑制されたmiR-301または過剰発現されたPTENは、ESCC細胞の移動と浸潤を抑制します。 miR-301の上昇またはPTENの低下は、ESCC細胞の移動と浸潤を誘発します
ESCC細胞の遊走および浸潤能力は、トランスウェルアッセイを使用して評価されました。結果は、KYSE-30細胞株(図4a、b)では、miR-301の阻害またはPTENの過剰発現によって細胞遊走および浸潤能力が抑制されたことを示唆しました。細胞遊走および浸潤能力におけるmiR-301阻害剤の抑制的役割は、si-PTENによって逆転されました。 Eca109細胞株(図4c、d)では、miR-301ミミックまたはsi-PTENのトランスフェクション後に細胞遊走および浸潤能力が促進されました。過剰発現したPTENは、miR-301ミミックが細胞の遊走および浸潤能力に与える影響を逆転させました。上記の発見は、ESCC細胞の移動と浸潤がmiR-301抑制またはPTEN上昇によって阻害されたのに対し、miR-301アップレギュレーションまたはPTENダウンレギュレーションによって促進されたことを意味しました。
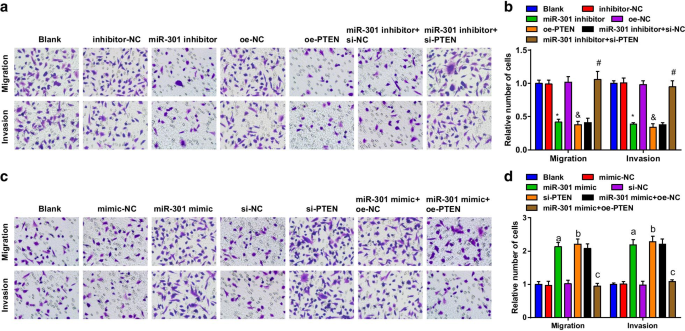
阻害されたmiR-301または過剰発現されたPTENは、ESCC細胞の移動と浸潤を抑制します。 miR-301の上昇またはPTENの低下は、ESCC細胞の移動と浸潤を促進します。 a miR-310のダウンレギュレーションまたはPTENのアップレギュレーション後にトランスウェルアッセイを使用して評価されたトランスフェクトされたKYSE-30細胞の遊走および浸潤能力。 b miR-310のダウンレギュレーションまたはPTENのアップレギュレーション後のトランスウェルアッセイによるKYSE-30細胞の遊走および浸潤の統計結果。 c miR-310のアップレギュレーションまたはPTENのダウンレギュレーション後にトランスウェルアッセイを使用して評価されたグループ間のEca109細胞の移動および浸潤能力。 d TranswellアッセイmiR-310のアップレギュレーションまたはPTENのダウンレギュレーションによるEca109細胞の遊走および浸潤の統計結果。 * P <0.05対阻害剤-NCグループ; & P <0.05対oe-NCグループ; # P <0.05対miR-301阻害剤+ si-NCグループ; P <0.05対mimic-NCグループ。 b P <0.05対si-NCグループ; c P <0.05対miR-301ミミック+ oe-NCグループ、 N =3.データは平均±標準偏差として表され、ANOVAは複数のグループ間の比較に使用されました
miR-301の阻害またはPTENの過剰発現は、細胞周期の停止とESCC細胞のアポトーシスを誘導します。 miR-301の上昇またはPTENの低下は、細胞周期の停止とESCC細胞のアポトーシスを抑制します
フローサイトメトリーを使用して、トランスフェクション後の細胞の細胞周期遷移とアポトーシスを検出しました。その結果、KYSE-30細胞株(図5a–d)では、miR-301阻害剤またはoe-PTENのトランスフェクションがアポトーシス率を促進することが示されました。 G0 / G1期では細胞が増加し、S期およびG2 / M期では細胞が減少しました。 miR-301阻害剤によって誘導されるアポトーシスと細胞周期停止の変化は、si-PTENによって逆転させることができます。
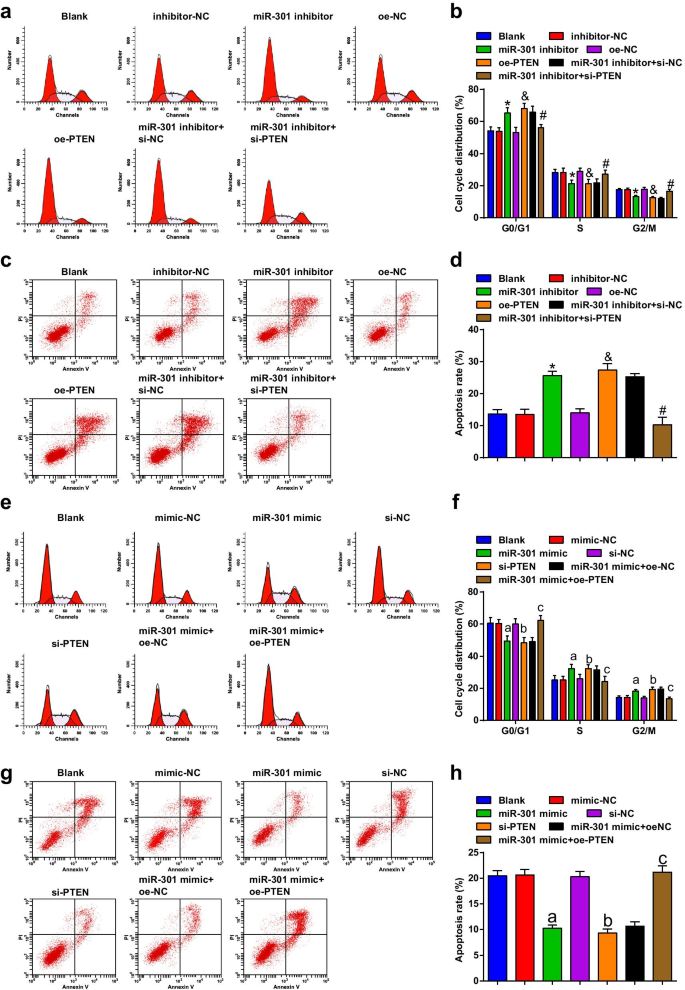
阻害されたmiR-301または過剰発現されたPTENは、ESCC細胞の細胞周期停止とアポトーシスを誘導します。 miR-301の上昇またはPTENの低下は、ESCC細胞の細胞周期停止とアポトーシスを抑制します。 a 各グループのKYSE-30細胞の細胞周期分布は、miR-310のダウンレギュレーションまたはPTENのアップレギュレーション後にフローサイトメトリーによって検出されました。 b miR-310ダウンレギュレーションまたはPTENアップレギュレーション後のフローサイトメトリーにおけるKYSE-30細胞のG0 / G1、SおよびG2 / GM期のパーセンテージの統計結果。 c KYSE-30細胞のアポトーシスは、miR-310のダウンレギュレーションまたはPTENのアップレギュレーション後にフローサイトメトリーによって検出されました。 d miR-310のダウンレギュレーションまたはPTENのアップレギュレーション後にフローサイトメトリーを使用して検出されたトランスフェクトされたKYSE-30細胞のアポトーシス率。 e 各グループのEca109細胞の細胞周期分布は、フローサイトメトリーmiR-310のアップレギュレーションまたはPTENのダウンレギュレーションによって検出されました。 f miR-310アップレギュレーションまたはPTENダウンレギュレーション後のフローサイトメトリーにおけるEca109細胞のG0 / G1、SおよびG2 / GM期のパーセンテージの統計結果。 g Eca109細胞のアポトーシスは、miR-310のアップレギュレーションまたはPTENのダウンレギュレーション後にフローサイトメトリーによって検出されました。 h miR-310のアップレギュレーションまたはPTENのダウンレギュレーション後にフローサイトメトリーを使用して検出されたトランスフェクトされたEca109細胞のアポトーシス率。 * P <0.05対阻害剤-NCグループ; & P <0.05対oe-NCグループ; # P <0.05対miR-301阻害剤+ si-NCグループ; P <0.05対mimic-NCグループ。 b P <0.05対si-NCグループ; c P <0.05対miR-301ミミック+ oe-NCグループ、 N =3.データは平均±標準偏差として表され、ANOVAは複数のグループ間の比較に使用されました
フローサイトメトリーの結果によると、Eca109細胞株(図5e–h)では、miR-301ミミックまたはsi-PTENのトランスフェクションがアポトーシス率を阻害し、G0 / G1期の細胞を減少させ、 S期およびG2 / M期; PTENの過剰発現は、Eca109細胞のアポトーシス率と細胞周期停止に対するmiR-301ミミックの効果を逆転させました。これらの結果から、miR-301のダウンレギュレーションまたはPTENのアップレギュレーションは、ESCC細胞の細胞周期の移行とアポトーシスを促進し、miR-301の阻害またはPTENのサイレンシングは反対の効果を発揮すると結論付けました。
抑制されたmiR-301または過剰発現されたPTENは、ESCCでinvivoで腫瘍の成長と血管新生を抑制します。 miR-301の上昇またはPTENの低下は、ESCCでinvivoで腫瘍の成長と血管新生を増加させます
ヌードマウスにおけるESCC腫瘍の成長と変化が各グループで観察された。腫瘍の成長を評価した結果、KYSE-30細胞株(図6a–e)では、miR-301阻害剤またはoe-PTENを注射したヌードマウスが腫瘍の体積と重量を減少させたことが示唆されました。腫瘍増殖におけるmiR-301阻害剤の抑制的役割は、si-PTENによって廃止されました。 Eca109細胞株(図6f–j)では、miR-301ミミックまたはsi-PTENを注射したヌードマウスの腫瘍体積と腫瘍重量が進行しました。 PTENの過剰発現は、腫瘍増殖に対するmiR-301模倣物の効果を逆転させました。一方、ヌードマウスの異種移植片におけるCD34の発現は、免疫組織化学的染色を使用して評価され、結果は、KYSE-30異種移植片では、miR-301のダウンレギュレーションまたはPTENのアップレギュレーション後にMVDが抑制されたことを示しました。沈黙したPTENは、MVDに対するmiR-301阻害の影響を逆転させました。 Eca109異種移植片では、miR-301のアップレギュレーションまたはPTENのダウンレギュレーション後にMVDが増加しました。上方制御されたmiR-301によって誘発されたMVDの増強は、過剰発現されたPTENによって無効にされる可能性があります。これらのデータは、miR-301の阻害またはPTENの過剰発現が、ESCCの腫瘍増殖と血管新生を抑制し、miR-301の上昇またはPTENのサイレンシングが逆効果をもたらしたことを示しています。

Inhibited miR-301 or overexpressed PTEN restrains tumor growth in ESCC; elevated miR-301 or reduced PTEN increases tumor growth in ESCC. a Representative figures for the tumor growth observed by subcutaneous tumorigenesis in nude mice after KYSE-30 cells were transfected; b–d changes of tumor volume of each group after KYSE-30 cells were transfected; e changes of tumor weight of each group after KYSE-30 cells were transfected; f representative figures for the tumor growth observed by subcutaneous tumorigenesis in nude mice after Eca109 cells were transfected; g–i changes of tumor volume of each group after Eca109 cells were transfected; j changes of tumor weight of each group after Eca109 cells were transfected. * P < 0.05 versus the inhibitor-NC group; &P < 0.05 versus the oe-NC group; # P < 0.05 versus the miR-301 inhibitors + si-NC group; P < 0.05 versus the mimic-NC group; b P < 0.05 versus the si-NC group; c P < 0.05 versus the miR-301 mimic + oe-NC group, n = 3 mice. Data were expressed as mean ± standard deviation, and ANOVA was used for comparisons among multiple groups

Inhibited miR-301 or overexpressed PTEN restrains angiogenesis in ESCC; elevated miR-301 or reduced PTEN increases angiogenesis in ESCC. a Representative images of tumor tissues observed by immunohistochemical staining in nude mice after KYSE-30 cells were transfected; b comparisons of MVD of KYSE-30 in tumor tissues among the groups; c representative images of tumor tissues observed by immunohistochemical staining in nude mice after Eca109 cells were transfected; d comparisons of MVD of Eca109 in tumor tissues among the groups *P < 0.05 versus the inhibitor-NC group; &P < 0.05 versus the oe-NC group; # P < 0.05 versus the miR-301 inhibitors + si-NC group; P < 0.05 versus the mimic-NC group; b P < 0.05 versus the si-NC group; c P < 0.05 versus the miR-301 mimic + oe-NC group, n = 3 mice. Data were expressed as mean ± standard deviation, and ANOVA was used for comparisons among multiple groups
ディスカッション
EC is a kind of invasive malignancy in the gastrointestinal tract [16]. As the major type of EC, ESCC is a malignant tumor occurring in esophageal epithelial cells [17]. The miRNAs, known as small non-coding RNAs, have been demonstrated to function as a significant roles in leading molecules in the silencing of RNA [18]. Our research was designed to explore the effects of miR-301 and its target gene PTEN on ESCC progression, and we have found that the inhibited miR-301 could suppress angiogenesis and cell growth in ESCC by elevating PTEN.
MiR-301 expression was assessed, and we found that miR-301 was highly expressed in ESCC cells in comparison with HEEC, and the higher expression of miR-301 has also been found in ESCC tissues in contrast to the adjacent normal tissues. Similar to this result, Li et al. have identified that miR-301 presented high expression in myocardial infarction tissues [19]. In addition, we have elucidated that PTEN was targeted by miR-301, and the target relation has been pointed out by an extant literature [20]. We have also discovered that PTEN, which has been affirmed to be targeted by miR-301, was downregulated in both ESCC tissues and cells. Similarly, a previous research has unearthed that PTEN was poorly expressed in ESCC compared with non-tumor esophageal epithelial tissue [21]. Furthermore, Ma et al. have illuminated that PTEN expression was degraded in Eca109 cell line [22], which has also been selected for a series of experiments in this research. These studies provide evidence for the high expression of miR-301 and low expression of PTEN in ESCC.
Another important outcome in this research indicated that the inhibited miR-301 could repress the colony formation ability as well as the cell proliferation of ESCC cells via enhancing the PTEN expression, and elevated miR-301 or reduced PTEN had contrary effects. Similarly, Han et al. have elucidated that the downregulation of miR-301 mediated by luteolin has the ability to restrain the cell proliferation in prostate cancer [6]. A recent literature has revealed that the overexpression of PTEN suppresses the proliferation of pancreatic cancer cells [23], and a same result has been summarized in a study focusing on prostate cancer [24]. Besides, we have also unearthed that the downregulation of miR-301 or the elevation of PTEN could inhibit migration and invasion of ESCC cells, and elevated miR-301 or reduced PTEN exhibited the opposite trends. In accordance with this outcome, Shi et al. have supported that inhibited miR-301 attenuated migration and invasion of breast cancer cells [10], and it has been reported that the migration and invasion of ESCC cells could be repressed by the inhibition of miR-130b and the elevation of PTEN [25]. These publications helped verifying the oncogenic role of miR-301 and tumor-repressive effect of PTEN in diverse human cancers. Another result in our research was that inhibited miR-301 overexpressed PTEN to promote cell apoptosis and induce cell cycle arrest at the G0/G1 phase in ESCC cells, and elevated miR-301 or reduced PTEN had the inverse results. Similarly, it has been uncovered by a recent literature that activated PTEN induces cell cycle arrest and apoptosis in ESCC [26]. Consistently, Tian et al. have found in their study that the elevation of PTEN inhibited the angiogenesis by reducing the expression of vascular endothelial growth factor in hepatocellular carcinoma [27]. Based on the above data, the roles of miR-301 and PTEN in cell apoptosis and angiogenesis in diverse diseases were further confirmed. Consequently, we concluded that the downregulation of miR-301 could restrain the tumor growth in ESCC through the high expression of PTEN, and the similar conclusion has also been unveiled in breast cancer [10] and prostate cancer [28]. On the contrary, miR-301 elevation or PTEN reduction induced the tumor growth in ESCC. It could be concluded that miR-301 and PTEN participated in the in vivo cancer cell growth.
Conclusion
In this study, we have shown that the repression of miR-301 prohibits angiogenesis, cell proliferation, migration and invasion but promotes apoptosis in ESCC cells by upregulating PTEN. This research may further the understanding on potential molecular mechanisms of ESCC and provide novel targets for ESCC treatment.
略語
- ESCC:
-
Esophageal squamous cell carcinoma
- PTEN:
-
Phosphatase and tensin homologue
- MVD:
-
Microvessel density
- EC:
-
Esophageal cancer
- miRNAs:
-
MicroRNAs
- LNM:
-
Lymph node metastasis
- UICC:
-
Union for International Cancer Control
- RT-qPCR:
-
Reverse transcription quantitative polymerase chain reaction
- 3′UTR:
-
3′-Untranslated region
- WT:
-
Wild type
- MUT:
-
Mutant type
- HRP:
-
西洋ワサビペルオキシダーゼ
- FBS:
-
ウシ胎児血清
- OE:
-
Overexpressed
- NC:
-
Negative control
- CCK-8:
-
Cell counting kit
- DMEM:
-
ダルベッコの改良イーグル培地
- PI:
-
ヨウ化プロピジウム
- FITC:
-
フルオレセインイソチオシアネート
- MVD:
-
Microvessel density
- ANOVA:
-
Analysis of variance
ナノマテリアル
- microRNA-342-5pのダウンレギュレーションまたはWnt3aのアップレギュレーションは、血管新生を阻害し、アテローム性動脈硬化症マウスのアテローム性動脈硬化症のプラーク安定性を維持します
- ICAをロードしたmPEG-ICAナノ粒子の調製とLPS誘発性H9c2細胞損傷の治療におけるそれらの応用
- ポリマーナノ構造上のU2OS細胞におけるアクチンと接着斑組織の分析
- 原子層堆積と水熱成長によって製造された抗菌性ポリアミド6-ZnO階層型ナノファイバー
- HT29およびSPEV細胞株に対するAuナノ粒子の影響のinvitro研究
- 6-メルカプトプリンとニューロン透過性ペプチドで修飾された金ナノ粒子によるSH-SY5Y細胞増殖の促進
- マゲマイトナノ粒子はナノザイムとして機能し、セイヨウアブラナの成長と非生物的ストレス耐性を改善します
- 丸い形の金ナノ粒子:シロイヌナズナの根の成長に対する粒子サイズと濃度の影響
- 直接メタノール燃料電池用の新しい陽極触媒のサポート:特性評価と単一セル性能
- InGaAs / InPコアシェルナノワイヤの自己シードMOCVD成長と劇的に増強されたフォトルミネッセンス
- 5Gと指数関数的なデータ増加の課題