


SOX2はlncRNACCAT1 / MicroRNA-185-3p / FOXP3軸を調節して、子宮頸がん幹細胞の増殖と自己再生に影響を与えます
要約
子宮頸がん(CC)における長鎖ノンコーディングRNA(lncRNA)の役割が示されています。性別決定領域Y-box2(SOX2)/ lncRNA結腸癌関連転写物-1(CCAT1)/ microRNA-185-3p(miR-185-3p)/フォークヘッドボックスタンパク質3(FOXP3 )CC幹細胞の増殖と自己複製能力について。 MiR-185-3p、SOX2、CCAT1およびFOXP3の発現は、CC組織および細胞でテストされました。 CC患者におけるSOX2 / CCAT1発現と臨床病理学的特徴との関係が検証された。 CD44 + で機能喪失と機能獲得の調査が行われました。 生物学的機能と自己複製能力について議論するHeLa細胞。最後に、SOX2、CCAT1、FOXP3、およびmiR-185-3p間の関係が検証されました。 miR-185-3pの発現は減少しましたが、SOX2、CCAT1、FOXP3の発現はCC組織と細胞で増加しました。 SOX2とCCAT1の発現は、腫瘍の大きさ、リンパ節転移、およびCCの婦人科と産科の国際連合に関連していた。 SOX2またはCCAT1をダウンレギュレートし、miR-185-3pをアップレギュレートすると、CD44 + のアポトーシス促進だけでなく、増殖、浸潤、遊走、細胞球数の阻害が生じました。 HeLa細胞。 SOX2はmiR-185-3pの発現に影響を与えるCCAT1に結合する可能性があり、FOXP3はmiR-185-3pの標的となりました。
はじめに
子宮頸がん(CC)は、2018年に世界中で推定57万人の症例と31万1000人の死亡を伴う女性の4番目の主要な死亡原因です[1]。この複雑な病気は、遺伝的影響やウイルス感染など、さまざまな要因に関与しています[2]。ヒトパピローマウイルスの同時検査とヒトパピローマウイルスワクチン接種の開発により、子宮頸部異形成と癌の早期診断手順により、CCの発生率、罹患率、死亡率が低下します[3]。初期のCC患者には、出産する手術、錐体生検、根治的子宮頸部摘出術、骨盤リンパ節郭清、骨盤放射線療法、近接照射療法などの手術が推奨されます[4]。進行したCC患者における転移または再発のために、予後は依然として不良である[5]。したがって、CCの治療を改善するための新しく効果的な予後マーカーと治療戦略を特定することは依然として緊急です。
性決定領域Y-box2(SOX2)は、転写因子SOXファミリーの重要なメンバーであり、主に胚性および成体幹細胞に発現し、腫瘍幹細胞にも発現します[6]。 SOX2がヘッジホッグシグナル伝達経路によってCCの放射線抵抗性を調節することが明らかになりました[7]。別の研究では、SOX2がCC細胞株の癌幹細胞の亜集団を維持するために重要であることが示されています[8]。長鎖ノンコーディングRNA(lncRNA)は、長さが200ヌクレオチドのRNA分子のクラスです[9]。 LncRNA結腸癌関連転写物-1(CCAT1)は、ヒト染色体8q24.21に位置し、結腸癌の遺伝子変異を引き起こす「ホットスポット」と見なされます[10]。ある研究では、CCAT1が細胞増殖とCCの浸潤を促進することが報告されています[11]。 Jia et al。によると、CCAT1はCC細胞の増殖、遊走、浸潤を劇的に促進します[12]。さらに、別の研究では、CCAT1がmicroRNA-185-3p(miR-185-3p)を減少させることにより、腸のバリアを破壊することにより、炎症性腸疾患の悪性度を高めることが明らかになっています[13]。 miRNAは、mRNAを減らし、翻訳を抑制することにより、遺伝子の発現を逆に制御することができます[14]。以前の研究でのinvitro実験は、miR-185-3pが鼻咽頭癌の放射線耐性を調節することを明らかにしました[15]。別の研究では、miR-185がinvivoおよびinvitroでの卵巣癌シスプラチン耐性に関与していることが示唆されています[16]。フォークヘッドボックスタンパク質3(FOXP3)は、FOXタンパク質ファミリーに属する転写因子であり、制御性T(Treg)細胞で最初に発見され、Treg細胞の維持とプロセスに重要な役割を果たします[17]。ある研究では、FOXP3がCCのリンパ管新生に関連していることが報告されています[18]。別の研究では、FOXP3のレベルが、婦人科および産科の国際連合の病期(FIGO期)およびCCの腫瘍サイズと劇的に関連していることが明らかになっています[19]。したがって、この研究では、CC幹細胞の増殖と自己複製能力に対するSOX2 / CCAT1 / miR-185-3p / FOXP3軸の影響を調べました。
材料と方法
参加するための倫理的承認と同意
人間が関与する実験は、ヘルシンキ宣言で表明された原則に従って実施されました。この研究は、吉林大学第一病院の倫理委員会によって承認されました。すべての参加者はインフォームドコンセントの文書に署名しました。
調査対象
2016年12月から2018年12月まで、CC組織および対応する隣接する正常組織の39例が、CC患者から採取され、液体窒素で保存されました。選択基準は次のとおりです。(1)患者は、経頸管および頸管の生検の病理学、頸部塗抹細胞診、頸部ヨウ素検査、膣鏡および頸椎切除によってCCとして確認されました。 (2)患者は、手術の2週間前に放射線療法と化学療法を受けていませんでした。除外基準は次のとおりです。(1)放射線療法または化学療法を受けている患者、(2)サンプルの収集に同意しなかった患者、(3)免疫系障害のある患者。
細胞の選択と培養
CC細胞株(SiHa、HeLa、CaSki、HCC94、C33A)およびヒト子宮頸部上皮不死化細胞株H8は、Shanghai Bioleaf Biotech Co.、Ltd。(上海、中国)から購入しました。 CC細胞SiHa、HeLa、およびHCC94は、10%ウシ胎児血清(FBS)を含む高グルコースダルベッコ改変イーグル培地(DMEM)で培養し、CaSki、C33A、およびH8細胞は、Roswell Park Memorial Institute(RPMI)-1640培地で培養しました。 %FBS(37℃および5%CO 2 )。細胞を剥がし、2日ごとに継代培養しました。
CC幹細胞の選別(CD44 + HeLa細胞)
CC幹細胞は、細胞浮遊培養によってCC細胞株HeLaから分離されました。 CC細胞HeLaは、無血清Iscoveの改良ダルベッコ培地(IMDM)により、超低接着性ペトリ皿で21日間培養され、培地は3〜5日ごとに半定量的に交換されました。一部の細胞を球体に懸濁し、HeLa球体形成細胞(SFC)を得た。球状細胞の特性が特定され、分析されました。細胞をトリプシンで剥離し、1×10 6 に調整しました 細胞/ mL。細胞にCD44抗体を加え、フローサイトメトリーで選別しました。 CD44が陽性のHeLa細胞はHeLa腫瘍幹細胞であり、CD44が陰性のHeLa細胞はHeLa非幹細胞でした。 CC幹細胞をDMEM / F12で培養し、20 ng / mLの塩基性線維芽細胞成長因子(bFGF)、20 ng / mLの表皮成長因子(EGF)、およびB27を添加しました。培地には1%のペニシリンとストレプトマイシンが含まれていました[20]。
細胞治療
CD44 + HeLa細胞にsh-SOX2、sh-SOX2ネガティブコントロール(NC)、sh-CCAT1、sh-CCAT1 NC、miR-185-3pミミック、ミミックNC、sh-CCAT1、miR-185-3p阻害剤をトランスフェクトしました。 sh-CCAT1および阻害剤NC。すべてのオリゴヌクレオチド配列は、GenePharma(上海、中国)から提供されました。トリプシンで分離した細胞を、3×10 6 の6ウェルプレートに播種しました。 細胞/ウェル。 60%のコンフルエンスに達したら、細胞を無血清培地に交換し、1時間インキュベートしました。トランスフェクションは、Lipofectamine 2000トランスフェクション試薬(Invitrogen、米国カリフォルニア州カールズバッド)によって促進されました。
逆転写定量的ポリメラーゼ連鎖反応(RT-qPCR)
組織および細胞内のRNAは、Trizol(Invitrogen)によって抽出されました。 RNA(1μg)は、モロニーマウス白血病ウイルスRTaseキット(Invitrogen)によってcDNAに逆転写されました。リアルタイムPCRシステムにcDNAを追加しました。プライマーはShanghaiSangon Biotechnology Co.、Ltd。(Shanghai、China)によって考案されました(表1)。 U6はmiR-185-3pのローディングコントロールであり、SOX2、CCAT1、FOXP3のグリセルアルデヒドリン酸デヒドロゲナーゼ(GAPDH)です。結果は2 -ΔΔCt によって分析されました メソッド。
<図>ウエスタンブロットアッセイ
細胞および組織中の総タンパク質が抽出されました。タンパク質濃度は、ビシンコニン酸キット(AmyJet Scientific、武漢、湖北省、中国)によって決定されました。タンパク質をローディングバッファーと混合し、5分間煮沸した後、氷浴と遠心分離を行いました。タンパク質を10%ドデシル硫酸ナトリウム-ポリアクリルアミドゲル電気泳動で処理し、メンブレンに転写しました。メンブレンを5%スキムミルクで1時間ブロックし、一次抗体SOX2(1:1000、Jiangsu Rui sitan Co.、Ltd。、江蘇、中国)、FOXP3(1:1000、Abcam Inc.、マサチューセッツ州ケンブリッジ)でプローブしました。 、USA)、GAPDH(1:1000、Cell Signaling Technology、マサチューセッツ州ビバリー、USA)、西洋ワサビペルオキシダーゼで標識された二次抗体で再プローブされ、保存フィルムで覆われ、露光されました。 Bio-rad Gel Doc EZイメージャー(Bio-rad、カリフォルニア、米国)が開発に採用されました。タンパク質画像はImageJ2xソフトウェアによって分析されました。
Cell Counting Kit(CCK)-8アッセイ
CCK-8アッセイは、キット(Beyotime、上海、中国)を使用して実装されました。セル(1×10 4 )を96ウェルプレートに播種し、インキュベートしました。 0、24、48、72時間培養し、細胞に10μL/ウェルのCCK-8溶液を加え、1時間孵化しました。光学密度の値は、450nmでMultiskanSpectrum全波長マイクロプレートリーダーを使用して決定されました。平均値を数えるために6つのウェルが取られました。細胞増殖曲線は、縦座標として時間、縦座標として相対的な細胞生存率でプロットされました。光学密度の値は細胞増殖を表しています。
フローサイトメトリー
セル(1×10 6 )を1500 r / minで遠心分離し、200μLの結合バッファーで懸濁し、5μLのヨウ化プロピジウム(PI)および5μLのアネキシンV-フルオレセインイソチオシアネート(FITC)と順番にインキュベートし、400μLのバッファーを添加しました。細胞アポトーシス率は、フローサイトメーター(BD Biosciences、NJ、USA)によって検証されました。
スクラッチテスト
トランスフェクトされた細胞を剥がし、細胞懸濁液に調製した。細胞懸濁液(1×10 6 細胞/ mL)を6ウェルプレートに播種し、80〜90%のコンフルエンスまで培養しました。 6ウェルプレートの横方向および縦方向の引っかき傷は、定規に沿った10μLの無菌ピペットチップによって均等に作成されました。元の培地を完全培地に交換し、細胞を継続的に培養した。細胞の移動は、0時間と72時間に顕微鏡で観察され、同じ場所で撮影されました。
トランスウェルアッセイ
トランスフェクトされた細胞をトリプシン処理し、1000 gで遠心分離し、DMEMですすいだ。完全に水和したトランスウェルチャンバーを、10%FBS-DMEM(600 mL /ウェル)を含む24ウェルプレートに配置しました。細胞懸濁液(5×10 5 細胞/ mL、200μL)をトランスウェル上部チャンバー(1:8マトリゲル80μLでコーティング)に添加し、500μLの20%FBS-DMEMを下部チャンバーに添加しました。細胞を24時間連続培養し、500μLの4%パラホルムアルデヒド(PFA)で固定し、0.1%クリスタルバイオレット染色液で染色しました。次に、上部チャンバーの表面と端の細胞を綿棒で拭き取った。 5つのフィールドをランダムに選択し、Nikon Eclipse TE2000-S顕微鏡(Nikon、日本)で細胞をカウントしました。
球体形成アッセイ
細胞は、200細胞/ウェルの無血清懸濁培地を含む低吸着6ウェルプレートに播種されました。 2週間後、Nikon Eclipse TE2000-S顕微鏡(Nikon)で細胞の球形成率を観察し、平均球数/播種細胞数×100%として球形成率を算出しました。
クロマチン免疫沈降(ChIP)アッセイ
ChIPアッセイは、ChIPキット(Upstate、NY、USA)を使用して操作しました。 SOX2(1:1000、Re-stem Biotech、江蘇省、中国)および通常のウサギIgG(12–370、ミリポア、米国)抗体を利用して、架橋タンパク質-DNA複合体を免疫沈降させました。 CD44 + HeLa細胞を1%PFAで固定し、インキュベートしてDNA-タンパク質架橋を生成しました。次に、超音波によってDNAを200〜300bpのクロマチンフラグメントに切断しました。沈殿したクロマチンDNAを回収し、RT-qPCRで分析しました。
RNAプルダウンアッセイ
ビオチン標識miR-185-3p野生型(WT)および変異型(MUT)プラスミド(50 nM)をCD44 + にトランスフェクトしました。 それぞれHeLa細胞。 48時間のトランスフェクション後、細胞を特定の細胞溶解物(Ambion、オースティン、テキサス、米国)で孵化しました。細胞溶解物(50 mL)をサブパッケージしました。残りのライセートを、RNaseフリーおよび酵母tRNA(Sigma)でプレコートしたM-280ストレプトアビジンビーズ(Sigma、セントルイス、ミズーリ州、米国)でハッチングしました。続いて、細胞を冷溶解物で2回、低塩緩衝液で3回、高塩緩衝液で1回洗浄した。拮抗的なmiR-185-3pプローブをNCとして設定しました。全RNAはTrizolによって抽出され、CCAT1レベルはRT-qPCRによってテストされました。
デュアルルシフェラーゼレポーター遺伝子アッセイ
CCAT1プロモーター領域上のSOX2の潜在的な結合部位E1およびE2は、https://jaspar.genereg.net/によって予測されました。 SOX2およびCCAT1結合部位E1を含むCCAT1プロモーター配列を合成し、CCAT1 3'UTR WT(E1-WT)およびCCAT1 3'UTR MUT(E1-MUT)ベクターを形成しました。ベクターはpmirGLO(Beyotime)にクローン化されました。その後、CCAT1-WT / pmirGLOまたはCCAT1-MUT / pmirGLOをsh-SOX2またはsh-SOX2NCでCD44 + にコトランスフェクトしました。 HeLa細胞をそれぞれ2日間培養した後、溶解しました。ルシフェラーゼ活性は、ルシフェラーゼ検出システム(タカラ、大連、中国)によってテストされました。
バイオインフォマティクスのウェブサイトは、CCAT1とmiR-185-3pの結合部位を予測および分析するために利用されました。 CCAT1とmiR-185-3pの結合部位は、デュアルルシフェラーゼレポーター遺伝子アッセイによって検証されました。 miR-185-3p結合部位を含むCCAT13'UTRを構成した。 CCAT1 3'UTRWTおよびCCAT13'UTR MUTが構築され、模倣物NCおよびmiR-185-3p模倣物でCD44 + にコトランスフェクトされました。 2日間のHeLa細胞。次に、細胞を溶解し、ルシフェラーゼ検出システム(タカラ)によってルシフェラーゼ活性を試験した。同じ方法を適用して、miR-185-3pとFOXP3の間のターゲティング関係を検証しました。
統計分析
すべてのデータは、SPSS 21.0ソフトウェア(IBM Corp. Armonk、NY、USA)によって評価されました。測定データは、平均±標準偏差として示されました。 t検定は、2つのグループ間の不一致と一元配置分散分析(ANOVA)に適用され、その後、グループ間の不一致についてテューキーの多重比較検定が行われました。分類変数は、フィッシャーの直接確率検定によって評価されました。 p 0.05未満の値は有意であると見なされました。
結果
miR-185-3pの発現は減少しますが、SOX2、CCAT1、FOXP3の発現はCC組織で増加し、SOX2とCCAT1の発現は腫瘍サイズ、リンパ節に関連しています転移(LNM)およびFIGOステージ
CC幹細胞の増殖および自己複製能力に対するSOX2 / CCAT1 / miR-185-3p / FOXP3軸の役割を検出した場合、CC組織および隣接する正常組織におけるmiR-185-3p、SOX2、CCAT1およびFOXP3の発現はRT-qPCRおよびウエスタンブロットアッセイによってテストされています。 (図1a–c)miR-185-3pの発現が減少し、CC組織ではSOX2、CCAT1、FOXP3の発現が増加したことが明らかになりました。
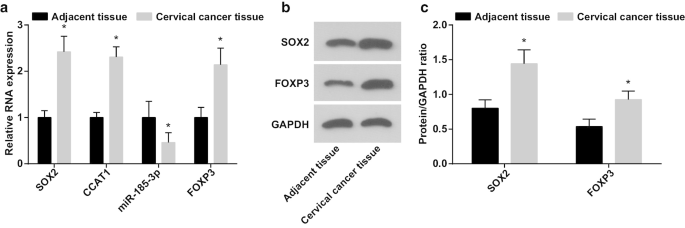
miR-185-3pの発現は減少しますが、CC組織ではSOX2、CCAT1、FOXP3の発現が増加します。 a CCおよび隣接する正常組織におけるSOX2mRNA、CCAT1、miR-185-3pおよびFOXP3mRNA発現の比較。 b CCおよび隣接する正常組織におけるSOX2およびFOXP3タンパク質発現のタンパク質バンド。 c CCおよび隣接する正常組織におけるSOX2およびFOXP3タンパク質発現の比較。 * p <0.05対隣接する正常組織。測定データは平均±標準偏差として表され、2つのグループ間の比較はt検定によって評価されました
SOX2 / CCAT1の発現とCCの臨床病理学的特徴との関係を分析しました(表2)。 CCで過剰発現したSOX2およびCCAT1は、腫瘍サイズ、LNMおよびFIGOステージと関連しており、SOX2およびCCAT1の発現は、CC患者の腫瘍サイズ、LNMおよび進行したFIGOステージが大きい患者で高かったことを示しています。
<図>miR-185-3p式は減少しますが、SOX2、CCAT1、およびFOXP3式はCD44で増加します + HeLa細胞
次に、ヒト子宮頸部上皮不死化細胞H8およびCC細胞株SiHa、CasKi、HeLa、HCC94、およびC33AにおけるmiR-185-3p、SOX2、CCAT1、およびFOXP3の発現をテストしました。 (図2a–c)miR-185-3pの発現は低下し、CC細胞株ではSOX2、CCAT1、FOXP3が上昇していることが示唆されました。その中で、HeLa細胞のSOX2 / CCAT1 / miR-185-3p / FOXP3はH8細胞との発現差が最も大きかった。したがって、HeLa細胞はスクリーニングされ、選別されました。 HeLa-SFCは細胞浮遊培養によって得られ、選別前後の細胞表面でのCD44発現はフローサイトメトリーによって同定されました。結果は、HeLa-SFCのCD44の陽性率が選別前よりも著しく高いことを強調し、CC幹細胞の選別が成功したことを示唆しています(図2d)。選別された幹細胞はCD44 + と名付けられました HeLa細胞。次に、miR-185-3p、SOX2、CCAT1、およびFOXP3の式をHeLaおよびCD44 + でテストしました。 HeLa細胞(図2e–g)。 CD44 + では、SOX2、CCAT1、FOXP3の発現がアップレギュレーションされ、miR-185-3pがダウンレギュレーションされたことが明らかになりました。 HeLa細胞。
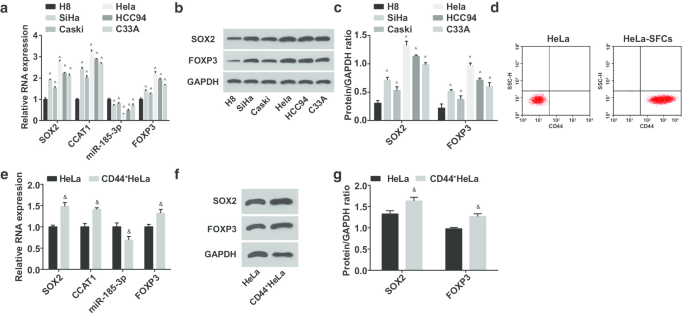
CD44 + では、miR-185-3pの発現が減少し、SOX2、CCAT1、FOXP3の発現が増加します。 HeLa細胞。 a H8およびCC細胞株におけるSOX2mRNA、CCAT1、miR-185-3pおよびFOXP3mRNA発現の比較。 b H8およびCC細胞株におけるSOX2およびFOXP3タンパク質発現のタンパク質バンド。 c H8およびCC細胞株におけるSOX2およびFOXP3タンパク質発現の比較。 d フローサイトメトリーによるソーティング前後のHeLa細胞におけるCD44発現率の検出。 e HeLa細胞とCD44 + 間のSOX2mRNA、CCAT1、miR-185-3p発現の比較 HeLa細胞。 f HeLa細胞およびCD44 + におけるSOX2タンパク質発現のタンパク質バンド HeLa細胞。 g HeLa細胞とCD44 + 間のSOX2タンパク質発現の比較 HeLa細胞。 ^ p <0.05対H8セル。 & p <0.05対HeLa細胞。測定データは平均±標準偏差として表され、2つのグループ間の比較はt検定によって評価され、複数のグループ間の比較は一元配置分散分析とそれに続くテューキーの事後検定によって評価されました
SOX2のダウンレギュレーションとCCAT1のダウンレギュレーションは、増殖、移動、侵入、球体形成率を低下させ、CD44のアポトーシスを増加させます + HeLa細胞
その後、SOX2とCCAT1はCD44 + でサイレンシングされました。 HeLa細胞は、CC幹細胞の増殖と自己複製への影響を調査します。 CCK-8アッセイ、フローサイトメトリー、スクラッチテスト、トランスウェルアッセイ、および球体形成実験によって検出され、CD44 + の増殖、移動、浸潤、および球体形成率が抑制され、アポトーシスが抑制されました。 HeLa細胞は、SOX2とCCAT1の阻害によって促進されました(図3a–i)。 SOX2またはCCAT1をサイレンシングすると、CC幹細胞の増殖と自己複製が阻害されたことが示されました。
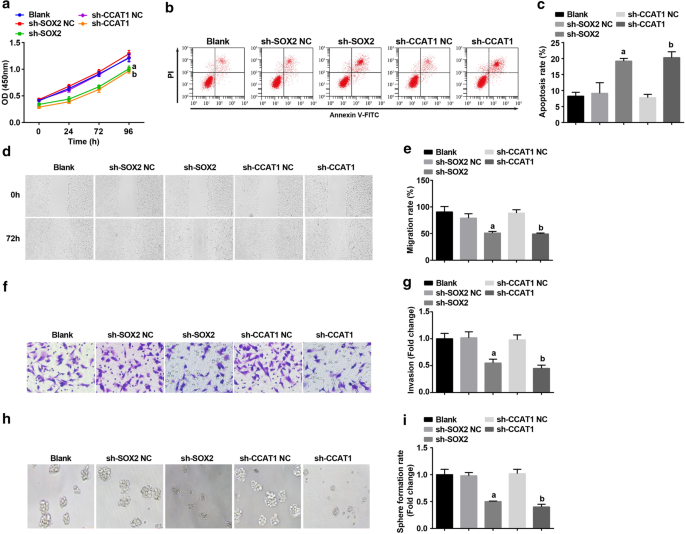
SOX2をダウンレギュレートし、CCAT1をダウンレギュレートすると、増殖、移動、浸潤、球体形成率が低下し、CD44 + のアポトーシスが増加します。 HeLa細胞。 a CCK-8アッセイでは、sh-CCAT1またはsh-SOX2で処理した細胞の細胞増殖曲線をテストしました。 b フローサイトメトリーは、sh-CCAT1またはsh-SOX2で処理された細胞の細胞アポトーシスを検出しました。 c sh-CCAT1またはsh-SOX2で処理した細胞の細胞アポトーシス率の比較。 d スクラッチテストでテストされたsh-CCAT1またはsh-SOX2で処理された細胞の細胞移動。 e sh-CCAT1またはsh-SOX2で処理された細胞における細胞遊走の比較。 f Transwellアッセイによるsh-CCAT1またはsh-SOX2で処理された細胞における細胞の浸潤能力の検出。 g sh-CCAT1またはsh-SOX2で処理した細胞の浸潤能の比較。 h 球体形成実験では、sh-CCAT1またはsh-SOX2で処理した細胞の自己複製能力をテストしました。 i sh-CCAT1またはsh-SOX2で処理した細胞の球体形成率の比較。 a p <0.05対sh-SOX2NCグループ。 b p <0.05対sh-CCAT1NCグループ。測定データは平均±標準偏差として表され、複数のグループ間の比較は一元配置分散分析とそれに続くテューキーの事後検定によって評価されました
枯渇したmiR-185-3pは、CD44におけるCCAT1のダウンレギュレーションの役割を逆転させます + HeLa細胞
次に、miR-185-3pがCC幹細胞の増殖と自己複製を調節するCCAT1のプロセスに関与しているかどうかを調べました。 CD44 + HeLaはmiR-185-3pミミックでトランスフェクトされたか、sh-CCAT1とmiR-185-3p阻害剤でコトランスフェクトされました。結果は、miR-185-3pのアップレギュレーションが、CD44 + のアポトーシス率を高めながら、増殖、移動、浸潤、および球体形成率を著しく低下させることを示しました。 HeLa細胞。 miR-185-3p阻害剤で処理された細胞は、CD44 + の増殖、遊走、浸潤、アポトーシス、および細胞球形成におけるダウンレギュレーションされたCCAT1の役割を逆転させる可能性があります。 HeLa細胞(図4a–i)。
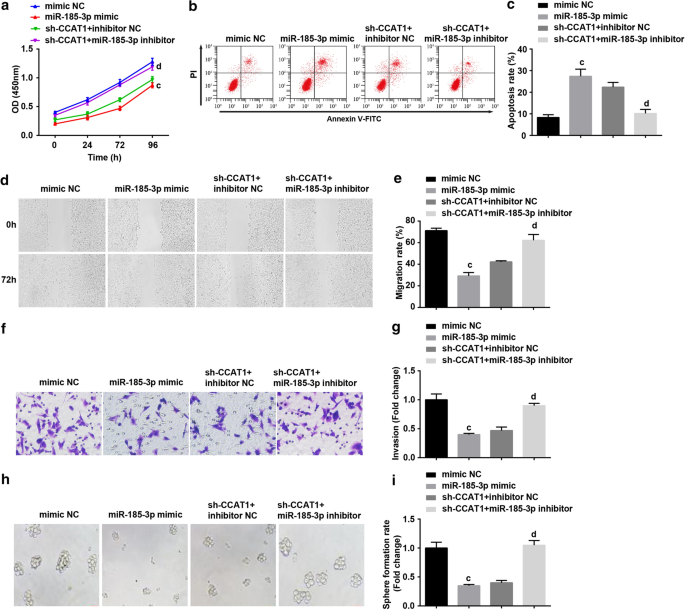
過剰発現したmiR-185-3pは、増殖、遊走、浸潤、球体形成率を抑制し、CD44 + のアポトーシスを増加させます。 HeLa細胞。 a CCK-8アッセイでは、miR-185-3pミミックで処理した細胞の細胞増殖曲線をテストしました。 b フローサイトメトリーは、miR-185-3pミミックで処理された細胞の細胞アポトーシスを検出しました。 c miR-185-3pミミックで処理した細胞の細胞アポトーシス率の比較。 d スクラッチテストでテストされたmiR-185-3pミミックで処理された細胞の細胞移動。 e miR-185-3pミミックで処理された細胞における細胞遊走の比較。 f TranswellアッセイによるmiR-185-3pミミックで処理された細胞の浸潤能力の検出。 g miR-185-3pミミックで処理された細胞の浸潤能力の比較。 h 球体形成実験では、miR-185-3pミミックで処理した細胞の自己複製能力をテストしました。 i miR-185-3pミミックで処理した細胞における球体形成率の比較。 a p <0.05対sh-SOX2NCグループ。 b p <0.05対sh-CCAT1NCグループ。 c p <0.05対模倣NCグループ。 d p <0.05対sh-CCAT1 +阻害剤NCグループ。測定データは平均±標準偏差として表され、複数のグループ間の比較は一元配置分散分析とそれに続くテューキーの事後検定によって評価されました
低発現SOX2および低発現CCAT1は、CD44 + でFOXP3の発現を減少させ、miR-185-3pの発現を増加させます。 HeLa細胞
その後、CD44 + でのSOX2 / CCAT1 / miR-185-3p / FOXP3の発現を調べました。 sh-SOX2、sh-CCAT1、miR-185-3pのトランスフェクション後のHeLa細胞は、sh-CCAT1とmiR-185-3p阻害剤の模倣および同時トランスフェクションを行います。 SOX2、CCAT1およびFOXP3の発現は減少しましたが、miR-185-3pの発現はsh-SOX2で処理した細胞で上昇しました。 CCAT1とFOXP3の発現が減少し、miR-185-3pの発現がsh-CCAT1で処理された細胞で増強されました。 miR-185-3p mimicを導入した細胞では、miR-185-3pの発現が上昇し、FOXP3の発現が低下しました。 sh-CCAT1とmiR-185-3p阻害剤を連続的にトランスフェクトした細胞では、FOXP3の発現が上昇し、miR-185-3pの発現が低下しました(図5a–d)。
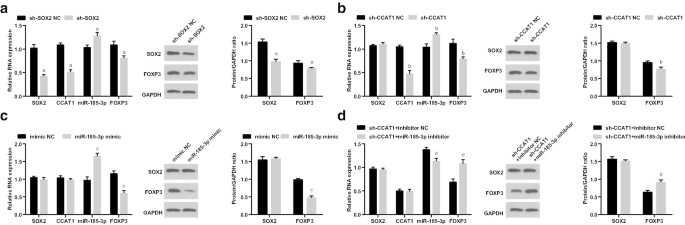
低発現のSOX2と低発現のCCAT1は、CD44 + でFOXP3の発現を減少させ、miR-185-3pの発現を増加させます。 HeLa細胞。 a sh-SOX2NCおよびsh-SOX2グループでのSOX2 / CCAT1 / miR-185-3p / FOXP3の発現。 b sh-CCAT1NCおよびsh-CCAT1グループでのSOX2 / CCAT1 / miR-185-3p / FOXP3の発現。 c 模倣NCおよびmiR-185-3p模倣グループにおけるSOX2 / CCAT1 / miR-185-3p / FOXP3の発現。 d sh-CCAT1 +阻害剤NCおよびsh-CCAT1 + miR-185-3p阻害剤グループにおけるSOX2 / CCAT1 / miR-185-3p / FOXP3の発現。 a p <0.05対sh-SOX2NCグループ。 b p <0.05対sh-CCAT1NCグループ。 c p <0.05対模倣NCグループ。 d p <0.05対sh-CCAT1 +阻害剤NCグループ。測定データは平均±標準偏差として表され、複数のグループ間の比較は一元配置分散分析とそれに続くテューキーの事後検定によって評価されました
SOX2はmiR-185-3pの発現に影響を与えるCCAT1に結合し、FOXP3はmiR-185-3pのターゲット遺伝子です
CCAT1プロモーター領域転写因子の潜在的な結合部位はhttps://jaspar.genereg.net/Webサイトによって予測および分析され、SOX2およびCCAT1がCCAT1プロモーター領域に潜在的な結合部位を持っていることが示されました(図6a)。 ChIP-qPCRは次のように報告しました(図6b):IgGグループとは対照的に、E1結合部位のSOX2グループではより多くのCCAT1プロモーターフラグメントが濃縮されており、SOX2がE1サイトのCCAT1プロモーターに結合し、SOX2がCCAT1の規制に関与しています。デュアルルシフェラーゼレポーター遺伝子アッセイは、次のことを示しました(図6c):ルシフェラーゼ活性はsh-SOX2とE1-WTでコトランスフェクトされた細胞で抑制され、SOX2がCCAT1に結合できることを示しています。
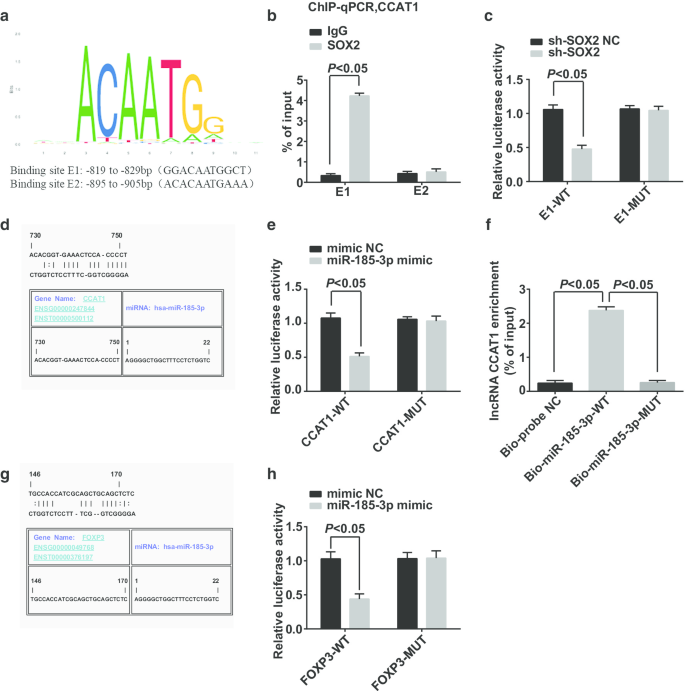
SOX2はmiR-185-3pの発現に影響を与えるCCAT1に結合でき、FOXP3はmiR-185-3pの標的遺伝子です。 a バイオインフォマティクスサイトによるSOX2およびCCAT1プロモーター領域の結合部位の予測。 b ChIP-qPCR実験により、SOX2とCCAT1の間の結合関係が検証されました。 c デュアルルシフェラーゼレポーター遺伝子アッセイによって検証されたSOX2とCCAT1の結合部位。 d バイオインフォマティクスサイトによるCCAT1およびmiR-185-3pの結合部位の予測。 e デュアルルシフェラーゼレポーター遺伝子アッセイによるCCAT1とmiR-185-3pの結合の検証。 f RNAプルダウンアッセイによって検証された細胞におけるCCAT1とmiR-185-3pの間の結合関係。 g バイオインフォマティクスのウェブサイトによるmiR-185-3pとFOXP3間のターゲティング関係の予測。 h デュアルルシフェラーゼレポーター遺伝子アッセイによるmiR-185-3pとFOXP3間のターゲティング関係の特定。測定データは平均±標準偏差として表され、2つのグループ間の比較はt検定によって評価されました
ジェファーソンのウェブサイトは、CCAT1がmiR-185-3pに結合する可能性があると予測しました(図6d)。デュアルルシフェラーゼレポーター遺伝子アッセイは、miR-185-3pミミックとCCAT1-WTを導入した細胞でルシフェラーゼ活性が低下したことを報告し(図6e)、miR-185-3pがCCAT1に結合できることを示唆しています。 RNAプルダウンアッセイを利用して、CCAT1がmiR-185-3pと結合できるかどうかを検証しました。その結果、(図6f)Bio-miR-185-3p-WTで処理した細胞のCCAT1の濃縮レベルが著しく増加したのに対し、Bio-miR-185-3p-MUTで処理した細胞のCCAT1の濃縮レベルは大きな違いはありません。この結果は、CCAT1がmiR-185-3pを吸着し、それによってmiR-185-3pの発現に影響を与える可能性があることを示しています。
miR-185-3pとFOXP3の間のターゲットの関係は、ジェファーソンのWebサイトによって予測されました(図6g)。デュアルルシフェラーゼレポーター遺伝子アッセイにより、FOXP3-WTとmiR-185-3pミミックをCD44 + にコトランスフェクトした後、細胞の相対的なルシフェラーゼ活性が劇的に低下することが確認されました(図6h)。 HeLa細胞は、miR-185-3pミミックでコトランスフェクトされたFOXP3-MUTは細胞の相対的なルシフェラーゼ活性に影響を与えなかったが、miR-185-3pがFOXP3を標的としたことを示唆している。
ディスカッション
CCは、世界で4番目に多い女性の悪性腫瘍であり、乳がん、結腸がん、肺がんがそれに続きます[3]。 SOX2発現が陽性のCC細胞は癌幹細胞の特徴を示すことが報告されています[21]。ある研究では、CCAT1はCCに関連する必須の発癌性lncRNAであり、CC細胞の成長と浸潤を促進する役割を果たしていることが報告されています[11]。別の研究では、miR-185-3pが鼻咽頭癌の放射線感受性を予測し、癌細胞の増殖とアポトーシスを調節する可能性があることが明らかになっています[22]。 FOXP3に対する循環自己抗体は、頸部病変の継続的な進行を反映しており、CCの早期予後の潜在的なバイオマーカーである可能性があることが報告されています[23]。現在の研究は、SOX2 / CCAT1 / miR-185-3p / FOXP3軸がCC幹細胞の増殖と自己複製能力にどのように影響するかを調査するために設計されました。
私たちの調査結果に基づいて、SOX2とCCAT1の発現は、腫瘍サイズ、LNM、および進行したFIGOに関連するCC組織および細胞で増加しました。機能的に証明された、SOX2とCCAT1のダウンレギュレーションは、増殖、遊走、浸潤、球体細胞数を減少させ、CC幹細胞のアポトーシスを増加させました。私たちの研究と同様に、SOX2の発現はCCで増加する傾向があります[24、25]。さらに、SOX2の発現は癌幹細胞に由来するCC細胞でもアップレギュレーションされます[26]。過剰発現したSOX2は、CCに限らず、いくつかの種類の癌患者の臨床病理学的特徴と関連していることが示唆されました。例えば、アップレギュレーションされたSOX2は、FIGOI-IIで病期分類された子宮頸部扁平上皮癌患者に見られることが示唆された[27]。さらに、SOX2の発現は口腔扁平上皮癌のLNMに関連しています[28]。癌の進行に対するSOX2の分子機能に関しては、ダウンレギュレーションされたSOX-2が子宮頸部扁平上皮癌の細胞移動と浸潤を抑制することを示す観察研究があります[29]。一方、別の研究では、アップレギュレーションされたSOX2が、対照細胞よりもinvitroおよびinvivoでCC細胞のクローン原性、増殖、腫瘍形成性を増強するという証拠が示されています[30]。
SOX2とCCAT1の間の調節関係に関して、既存の研究は、SOX2をサイレンシングするとCCAT1mRNAレベルが著しく低下することを示しています[31]。癌におけるCCAT1の役割に関して、ある研究では、CCAT1の発現が隣接する正常組織と比較してCC組織で著しく上昇していることが示されています[11、12]。注目すべきことに、CCでのCCAT1の過剰発現は、腫瘍のサイズと正の関係があります[12]。癌細胞の活動におけるCCAT1の役割に関して、過剰発現したCCAT1がCC細胞の増殖、コロニー形成、および浸潤を加速することを強調する研究があります[11]。興味深いことに、以前の研究では、CCAT1をノックダウンすると、細胞の生存能力、浸潤性、遊走性が低下することが示されています[12]。とにかく、他の癌におけるSOX2とCCAT1の機能的効果はCCのそれと同様でした。
その後、私たちの研究により、CCAT1がmiR-185-3pに結合できることが明らかになりました。これは、CCでダウンレギュレーションされたCCAT1であり、miR-185-3pを過剰発現させると、CC幹細胞の増殖と自己複製能力が抑制されます。 CCAT1とmiR-185-3pは負の相関があると報告されています[13]。さらに、結果は、放射線抵抗性鼻咽頭癌の症例でmiR-185-3p発現の減少が見られることを報告しました[22]。癌細胞の攻撃性におけるmiR-185-3pの抑制機能に関して、研究により、miR-185-3pのアップレギュレーションが鼻咽頭癌細胞の浸潤性および転移性を抑制することが明らかになりました[32]。さらに、Zou etal。復元されたmiR-185が乳がん細胞の成長と浸潤を抑制することを示唆しています[33]。 miR-185のアップレギュレーションがinvitroでの非小細胞肺癌細胞の増殖、浸潤およびコロニー形成能力を低下させることを発見した記事があります[34]。非小細胞肺癌細胞でのmiR-185の過剰発現により、in vitroでの細胞増殖、浸潤、遊走、およびinvivoでの腫瘍増殖が抑制されることが示されています[35]。これらの研究から、本研究におけるmiR-185-3pの抗腫瘍の役割は、以前の研究と一致していました。
先に進むために、miR-185-3pがCCで過剰発現している遺伝子であるFOXP3を標的にしてCC幹細胞の活性を調節していることを明らかにしました。実際、結腸癌におけるSOX2癌幹様細胞マーカーの調節因子であるFOXP3 [36]は、CCで調査されており、CC細胞でのアップレギュレーションを示しています[19] [37]。 FOXP3を上昇させると、腫瘍球の形成が促進され、非小細胞肺がん細胞の幹細胞性が刺激されることが証明されました[38]。
結論
まとめると、SOX2転写がCCAT1を活性化し、それによってmiR-185-3pを阻害し、FOXP3を調節して、CCを治療する潜在的な手段であるCC幹細胞の増殖と自己複製を促進できることを初めて調査しました。さらに、しかしながら、この現在の研究における制限は、計画された実験における比較的小さな試行サイズにまだ存在している。したがって、CCの治療の有効性を検出するために臨床研究がさらに実施される可能性があります。
データと資料の可用性
該当なし。
ナノマテリアル
- 卵巣癌細胞の高感度かつ迅速な検出のための柔軟なグラフェンベースのバイオセンサーのデモンストレーション
- 癌治療のためのナノ粒子:現在の進歩と課題
- 癌用途のための細胞ベースのドラッグデリバリー
- ナノテクニックは癌幹細胞を不活性化する
- 色素増感太陽電池の光学的および電気的特性に及ぼすTiO2中の金ナノ粒子分布の影響
- 酸化グラフェンハイブリダイズしたnHAC / PLGA足場はMC3T3-E1細胞の増殖を促進します
- 二層/三層ブロードバンドSiO2反射防止膜の断面形態に関するTEMおよびSTEM研究
- ポリオール媒介プロセスによるZnOナノクリップの製造と特性評価
- 小胞および細胞の周波数変調波誘電泳動:クロスオーバー周波数での周期的Uターン
- ペロブスカイト太陽電池の変換効率に及ぼすTiO2コンパクト層に埋め込まれた異なるサイズと濃度のAgナノ粒子の影響
- パンチングプロセスの成功と安全性に影響を与える要因