


新鮮および老化した酸化亜鉛ナノ粒子によって誘発される細胞毒性におけるアポトーシス経路の役割
要約
酸化亜鉛ナノ粒子(ZnO NP)は、産業、商業製品、医療分野など、幅広い用途で使用されています。 ZnO NPの毒性に関する多数の機構的研究が、元の(新鮮な)NPで実施されました。ただし、変換された(老化した)ZnONPによって誘導される細胞毒性とその根底にあるメカニズムは不明なままです。ここでは、時間の経過とともに行われたZnO NPの物理化学的変換を観察し、続いて新鮮なNPと老化したNPの細胞毒性を評価しました。我々は、新鮮なZnONPがそれらの老化した対応物よりも高いアポトーシスレベルを誘導することを発見した。したがって、老化したZnO NPで処理されたヒト-ハムスターハイブリッド( A )からのRNAシーケンスデータ L )細胞は、p53、PI3k–Akt、FoXO、グルタチオン、ErbB、HIF-1、オキシトシン、およびJak-STATシグナル伝達経路が濃縮されているが、アポトーシス経路は濃縮されていないことを示しました。定量的PCRの結果から、 IL1B のmRNAレベルが大幅に高いことが明らかになりました。 および CD69 老化したZnONPおよび塩化亜鉛処理群と比較した新鮮なNP処理群で。上記の結果は、老化したZnO NPの細胞毒性が低いのは、アポトーシスを誘導する効力が低下していることに部分的に起因していることを示しています。老化したNPによって活性化される複数のシグナル経路の転写調節は、細胞の恒常性を構築するのに役立つ可能性があります。まとめると、私たちの調査結果は、ZnO NPの老化(環境変換)プロセスが毒性と生物学的影響に及ぼす影響を浮き彫りにしています。
はじめに
過去数十年にわたるナノテクノロジーの急速な発展に伴い、ナノ粒子(NP)は、産業、人間の日常生活、ナノ医療など、さまざまな分野で適用されてきました[1、2]。ナノテクノロジー消費者製品目録(CPI)は、健康(フィットネス)製品762、食品(飲料)72、ベビー製品23を含むナノ製品の数が2005年から2015年の間に30倍に増加したことを示しています[2]。消費者製品やさまざまな分野でのNPの適用の増加により、NPが環境に侵入する可能性が高まり、潜在的な悪影響に関する安全上の懸念が生じています。酸化亜鉛(ZnO)NPは、最も一般的に使用されているNPの1つであり、その世界の年間生産量は3400トン近くに達しています[3、4]。以前は生物学的に不活性であると考えられていたいくつかの物質は、ナノ粒子状態で有毒になる可能性があります。ますます多くの研究が、ZnONPが重大な毒性を誘発することによって哺乳類の細胞や動物に重大なリスクをもたらす可能性があることを明らかにしました[5,6,7]。
コーティング、表面機能化、酸化状態の変更などのさまざまな戦略を使用して、NPの物理的および化学的特性(細胞膜の溶解、凝集、摂動など)を変更することにより、NPの潜在的な毒性を軽減しています[8,9,10、 11]。 NPのこれらの変更はある程度毒性効果を弱めますが、NPの使用は、特に特定の曝露条件および環境下では、常に安全であるとは限りません[12、13、14]。実際、多くの種類のNPは安定しておらず、意図的または意図せずに自然環境に放出された後、「老化」または「環境変化」を起こす傾向があります[14、15、16、17]。近年、NPの環境変換プロセスを調査するために多くの作業が実施されました。ただし、「変換された(老化した)」NPの毒性効果に関する研究は、毒性メカニズムは言うまでもなく、まだ非常に限られています。
非持続性NPの典型的な代表として、ZnO NPは非常に高い反応性を持ち、環境に放出された後、または動物に摂取された後、物理的および化学的特性と発生状態に変化する傾向があり、毒性学的影響に大きな影響を与える可能性があります[17 、18]。たとえば、研究によると、ZnO NPの硫化プロセスにより、電荷、疎水性、凝集状態が変化し、ヒトの唾液、汗、気管支肺胞洗浄液に硫化物状態のNPが吸着することが示されています。さらに、ZnO NPによって吸着されたタンパク質は、通常その生物学的効果に影響を与える特別なタンパク質クラウンを形成します[19]。生理的溶液中のリン酸塩は、ZnONPを準安定ZnHPO 4 に変換する可能性があります およびZn 3 (PO 4 ) 2 約5〜10時間以内[20]。ヒトTリンパ球(37°C、10%FBSを含む細胞培養培地RPMI1640を24時間)へのin vitro曝露システムでのZnO NP(≤3μg/ mL)の完全な変換のプロセスを、放射光X-を使用して調査しました。光線吸収端構造分光法(XANES)[21]。上記の研究は、純粋な(新鮮な)ZnO NPの生物学的効果を評価するだけで、ZnONPの環境および健康リスクを過小評価していることを示唆しています。この問題に照らして、NPの老化と環境変化のプロセスを包括的に理解することが急務である[22]。
以前の調査では、超純水中で40〜120日間エージングされたZnO NPは、物理化学的変換を受けてZn 5 に変わることが明らかになりました。 (CO 3 ) 2 (OH) 6 、Zn(OH) 2 、およびZn 2+ [23]。興味深いことに、老化したZnO NPは、新鮮な対応物よりも低い細胞毒性を示しました[23]が、そのような種類の変動の毒性メカニズムは不明です。本研究では、新鮮なZnONPと老化したZnONPの間の異なる細胞毒性の根本的な理由を調査することに着手しました。 2つの異なる粒子サイズ(20 nmと90〜200 nm)のZnONPを体系的に適用しました。細胞毒性アッセイは、老化したZnO NPが、それらの新鮮な対応物よりも目立たない形態学的異常と比較的高い細胞生存率を誘発することを示した。 RNAシーケンスデータは、アポトーシス遺伝子が新鮮なZnO NP処理細胞で濃縮されているのに対し、これらの遺伝子は老化したZnONP処理による影響がはるかに少ないことを明らかにしました。さらに、老化したZnO NPに曝露された細胞は、切断されたカスパーゼ-3タンパク質のレベルの低下を示し、培養細胞でアポトーシスを誘発する新鮮なZnONPの効力が高いことをさらに示しています。以前の調査結果と組み合わせて、この研究は、老化したZnO NPの細胞毒性の低下は、細胞アポトーシスを誘発する能力の低下に起因することを示唆しました。
材料と方法
ナノ粒子と試薬
市販のZnOナノ粉末(ZnO NP)は、メーカーの報告による平均サイズ20 nm(99.5%純度、ほぼ球状)および90〜200 nm(99.9%純度、不規則な形態)で、Nanostructured&Amorphous Materials(Houston、TX)から購入しました。 )。特に明記されていない限り、この研究で使用されたすべての試薬と化学物質は、Sigma-Aldrich(上海、中国)から購入しました。
ナノ粒子の分散、エージング、および特性評価
乾燥ナノ粉末をミリ- Q に懸濁することにより、ZnO NPストック懸濁液(1 mg / mL)を調製しました。 H 2 O(ミリポア、18MΩcm)、オートクレーブ滅菌(120°C、30分)した後、25°Cで0〜60日間の自然老化期間保存します。 0日および60日の自然に変換されたZnONPは、それぞれ新鮮なNPおよび老化したNPとして指定されました。適切な分散を確保するために、新鮮な懸濁液と熟成した懸濁液を超音波浴で30分間超音波処理(100 W)した後、特性評価または細胞とのインキュベーションを行いました。新鮮なおよび老化したZnONPの形態、粒子サイズ、および凝集は、透過型電子顕微鏡法(TEM、JEOL JEM-2010、東京、日本)を使用して特徴づけられました。新鮮なおよび老化したZnONPの結晶構造は、本物の標準と比較することにより、パワーX線回折(XRD、PANalytical B. V.、上海、中国)を使用して決定されました。 ZnO NPの自然老化プロセスと特性評価の詳細は、以前に説明されています[23]。
ZnONPによる細胞培養と処理
A L 細胞株、 gly2A の融合によって形成されたヒト-ハムスターハイブリッド細胞の一種 この研究では、チャイニーズハムスター卵巣(CHO)とヒト線維芽細胞の変異体を使用しました。これらのハイブリッド細胞は、CHO-K1染色体の標準セットとヒト11番染色体の単一コピーを含み、ウシ胎児血清(8%、ニューヨーク州グランドアイランドのハイクローン)を添加したハムのF12培地(ニューヨーク州グランドアイランドのハイクローン)で培養されました。 )、ゲンタマイシン(25 g / mL)およびグリシン(2×10 –4 M)加湿した5%CO 2 中37°Cで インキュベーター[24]。新鮮なおよび老化したZnONPのストック懸濁液を、凝集を防ぐために30分間の超音波処理(100 W)で分散させた後、細胞を曝露するために細胞培養培地で適切な濃度に希釈しました。 NPを含まない細胞培養培地で維持された細胞は、各実験でコントロールとして使用されました。
細胞毒性を検出するためのアッセイ
A L 対数増殖期の細胞を35mmペトリ皿(6×10 4 )のスライドガラス上で培養しました。 細胞/皿)を刺激する前に24時間処理した後、1、5、10、12、15、および20 µg / mLの新鮮または老化したZnONPを含む2mLの培地で72時間処理します。処理時間の終了後、ライカDM4B顕微鏡(ライカ、ドイツ)を使用して細胞形態の画像を取得しました。 ZnCl 2 細胞毒性をZnONPと比較するための亜鉛イオンリファレンスとして含まれています。
細胞生存率を検出するために、細胞計数キット(CCK-8)(APExBIO、上海、中国)を使用しました。詳細は、 A L 細胞を96ウェルプレートに播種しました(4×10 3 細胞/ウェル)細胞培養培地で24時間処理し、さまざまな濃度のZnCl 2 を含む培地で処理 、それぞれ24時間、48時間、72時間の新鮮なおよび老化したZnONP。作業溶液の場合、ストック懸濁液から添加されたNPの量は、各ウェルの培地の総量の5%未満でした。処理時間の終了後、培地を吸引し、製造元の指示に従って、細胞を100 µLのCCK-8作業溶液とともに37°Cで2時間インキュベートしました。次に、Spectra Max M2蛍光リーダー(Molecular Devices、Wokingham、Berks、UK)を使用して、450nmで吸光度を記録しました。細胞生存率は、ウェル内の吸光度のパーセンテージとして計算され、NPの各濃度はコントロール細胞の吸光度(100%)に正規化されています。
RNA抽出、逆転写、定量PCR
A L 対数増殖期の細胞を直径35mmのペトリ皿(6×10 4 )に播種しました。 細胞/皿)細胞培養培地で24時間。次に、培地を12 µg / mLのZnCl 2 を含む2mLの培地に交換しました。 、72時間の新鮮で熟成したZnONP。処理時間の終了後、培地を吸引し、細胞をPBSで3回洗浄した。続いて、1 mLのTrizol試薬(Invitrogen、Carlsbad、CA、USA)を各ディッシュに追加し、製造元の指示に従ってトータルRNAを抽出しました。抽出後に得られたトータルRNAの濃度と純度は、Q5000UV-Vis分光光度計(Quawell、USA)を使用して定量化されました。定量後、TransGene RT-PCRキット(TransGene Biotech、北京、中国)を使用して逆転写を行い、製造元のプロトコルに従ってRNAテンプレートからcDNAを取得しました。得られたcDNAサンプルは、Q5000 UV-Vis分光光度計を使用して定量し、Roche RT-PCRシステム(Applied Biosystems)で蛍光色素としてSYBR-Green(TransGene Biotech、北京、中国)を使用して分析しました[25]。
グリセルアルデヒド-3-リン酸デヒドロゲナーゼ( Gapdh )をコードするハウスキーピング遺伝子 )Il-1αを評価するための内部統制として使用されました 、Il-1β 、カスパーゼ3 、 CD69 、 6月 および MT1 mRNAの発現。結果は、標的遺伝子と Gapdh の間の相対的な発現比として表されました。 。この研究で使用したプライマー配列を表1に示します。
<図>RNAシーケンシングデータ分析
A のトータルRNAサンプル L 対照群、老化したZnO NP処理群、およびZnCl 2 の細胞 治療群はBangFeiBioscience(北京、中国)によって配列決定されました。簡単に説明すると、 A のトータルRNA L 細胞は、等プロポナール沈殿まで、TRIZOLプロトコルに従って抽出されました。次に、RNAサンプルを抽出バッファーに再懸濁してからシーケンスを行いました。生カウントRNAシーケンスデータは、RパッケージDeseq2 [Eric1]を使用して分析されました。ベン図は、RパッケージVennDiagram [Eric1.2]によって生成されました。大幅に変更された遺伝子は、さらなる経路濃縮分析に使用されました。実験は3回の独立した複製で行われました。 rRNA遺伝子、ミトコンドリア遺伝子、および40bp未満で検出された遺伝子は分析から除外されました。
RNAシーケンシングデータ、リファレンスシリーズGSE97852、GSE60159、GSE39444は、Gene Expression Omnibus [Eric 2、3、4]から入手しました。 Gene Set Enrichment Analysisプロットは、パッケージfgsea [Eric 5]を使用してR(バージョン3.6.2)によって生成されました。 1.5倍の有意な変化と p を持つアポトーシス遺伝子 値<0.05をさらに分析するために使用しました。遺伝子ツリーを含むヒートマップは、Rパッケージ「ComplexHeatmap」[Eric6]によって生成されました。クラスタリング手法として平均リンケージを使用し、距離測定手法としてユークリダンを使用した。経路濃縮分析の前に、STRING2.0 [Eric7]を使用しました。
ウエスタンブロッティング
A L 対数増殖期の細胞を直径60mmのペトリ皿(1.5×10 5 )に播種しました。 細胞/皿)細胞培養培地で24時間。次に、培地を、12 µg / mLの新鮮または老化したZnONPを含む4mLの培地と24時間交換しました。曝露期間の終わりに、培地を吸引し、次に細胞をPBSで3回洗浄し、RIPA溶解緩衝液(Beyotime、中国)を用いて氷上で溶解して細胞タンパク質を収集した。等量の細胞タンパク質を12%SDS-PAGEゲルで分離し、ポリフッ化ビニリデン(PVDF)メンブレン(Roche、Swiss)に転写しました。簡単に説明すると、25°CでTBST中の5%脱脂乳で2時間ブロッキングした後、膜を適切な希釈率の1次抗体(製造元のプロトコルに従って)と4°Cで一晩インキュベートし、続いてHRP結合二次抗体とインキュベートしました。抗体(1:5000、Promega、マディソン、米国)、25°Cで2時間。最後に、強化化学発光(ECL)(BOSTER、中国)溶液を使用して免疫標識を検出しました。抗プロ/切断型カスパーゼ-3および抗アクチンの一次抗体は、それぞれCell SignalingTechnologyおよびImmunoWayから購入しました。
統計
統計分析は、少なくとも3回の独立した実験から得られた結果を基にまとめられました。すべてのデータは平均±標準偏差(SD)として提示され、一元配置分散分析(ANOVA)を使用して統計的に比較されました。すべてのプロットで p 値<0.05は*として示され、統計的に有意であると見なされました。
結果
ZnONPの特性評価
新鮮なZnONPと古いZnONPの詳細な物理化学的特性の違いを判断するために、まずTEMを使用してNPの形態を観察しました(図1A)。私たちの結果は、20nmの新鮮なZnONPはほぼ球形の結晶であり、90〜200nmの新鮮なZnONPは不規則な棒状/立方体の結晶であることを示しました。単一粒子サイズは、製造業者によって提供されたサイズと一致していた。明らかに、20 nmと90〜200nmの両方のZnONPは、超純水中で凝集する傾向がありました。また、元のNPの形状とサイズに関係なく、20 nmと90〜200nmの両方のZnONPの微細構造は、60日間エージングした後、透明な結晶構造からアモルファスまたはシート/針のような状態に劇的に変化しました。さらに、新鮮なNPと老化したNPの両方の結晶性と相純度は、CuKα線(λ)を使用したX線回折(XRD)を使用して決定されました。 =0.15418 nm)図1Bに示すように、25°Cでアプローチします。新鮮なZnONPのXRDパターンは、サンプルが結晶性ウルツ鉱構造で構成されており、特徴的な不純物ピークが特定されていないことを示しており、新鮮なNPの品質が高いことを示しています。老化したNPの場合、XRDパターンはZn 5 の新形成を示しました (CO 3 ) 2 (OH) 6 (カード番号00-011-0287)およびZn(OH) 2 (カード番号00-003-0888)固相、エージングプロセス中のZnO NP(20および90–200 nm)の化学変換を示します。
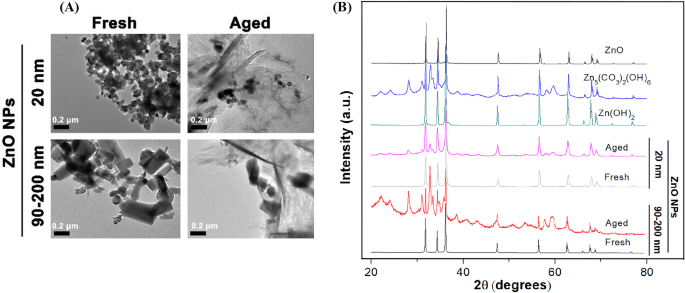
新鮮および老化したZnONPの物理化学的特性。 A Milli- Q における新鮮および老化したNP(100μg/ mL、20および90〜200 nm)の代表的な顕微鏡写真 低分解能TEMを使用した水、 B 新鮮なNP、老化したNP、ZnO、Zn(OH) 2 のXRDパターン およびZn 5 (CO 3 ) 2 (OH) 6 乾燥した形の参照
A の形態学的観察 L 新鮮で老化したZnONPに曝露された細胞
NPの治療は、invitroで細胞の形状または形態に顕著な変化をもたらします[26]。したがって、 A L 10および15µg / mLの新鮮または老化したZnONPに72時間曝露した細胞を、実体顕微鏡で検査しました。図2に示すように、対照群の細胞形態は正常なままでした。細胞はよく接着し、ほとんどが2時間以内に付着しました。ほとんどの細胞は紡錘形または多角形であり、いくつかの新しく分裂した細胞は、接着の過程でより透明な細胞質とより良い分散を示しました。 12μg/ mLの新鮮なZnONP(20 nmおよび90〜200 nm)で72時間処理すると、細胞の形態が大幅に変化しました。ほとんどの細胞は3〜5時間以内に付着しましたが、うまく広がることができず、一部の細胞は丸くなり、多角形を失いました。 ZnONPの濃度を15μg/ mLに上げると、処理された細胞は萎縮して付着できなくなり、10μg/ mLで処理された細胞よりも細胞生存率が大幅に低下したことが示唆されました。これらの結果は、新鮮なZnONPのLC100が72時間の処理でおそらく15μg/ mL未満であることを示しています。対照的に、20 nmおよび90〜200 nmの老化したNP処理グループ(15μg/ mL)の細胞形態は大きな影響を受けず、生き残った細胞のほとんどが付着して広がる可能性があり、死んだ細胞の半分未満が観察され、老化したZnONPは、新鮮なZnONPよりも細胞毒性がはるかに低いです。
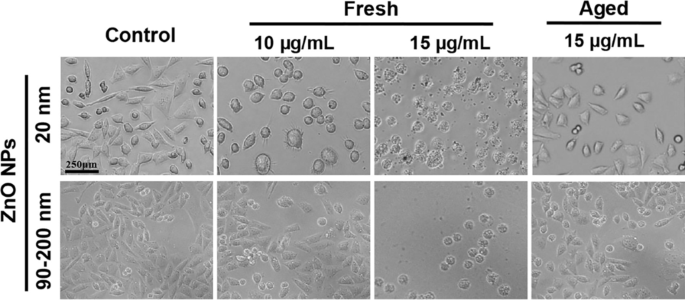
A の形態変化 L 補足のハムのF12培地で72時間新鮮または老化したZnONPに曝露した後の細胞、および曝露されていない細胞を対照群として使用しました。 A L 細胞の形態を光学顕微鏡で10×倍率で観察しました
老化したZnONPは、新鮮なNPよりも低い細胞毒性を誘発しました
新鮮なZnONPと老化したZnONPの細胞毒性の違いをさらに調査するために、CCK-8キットを使用して細胞の生存率を調べました。図3に示すように、インキュベーション A L 新鮮および老化したZnONP(0〜20μg / mL、20 nm、および90〜200 nmの範囲)の勾配用量を24時間、48時間、または72時間使用した細胞では、細胞生存率の用量依存的な低下が見られました。細胞をZnONPs≤10μg/ mLで処理しても、明らかな細胞毒性は観察されませんでした。新鮮なおよび老化したZnONPの投与量が12および15μg/ mLに上昇すると、細胞生存率は時間依存的な減少傾向を示しました。明らかに、老化したNP治療群の細胞生存率は、新鮮なNP治療群よりも有意に高かった。さらに、ZnCl 2 -処理はまた、用量および時間依存的に細胞生存率を低下させましたが、ZnCl 2 の細胞毒性は 新鮮なZnONPと老化したZnONPの両方よりもはるかに少なかった。
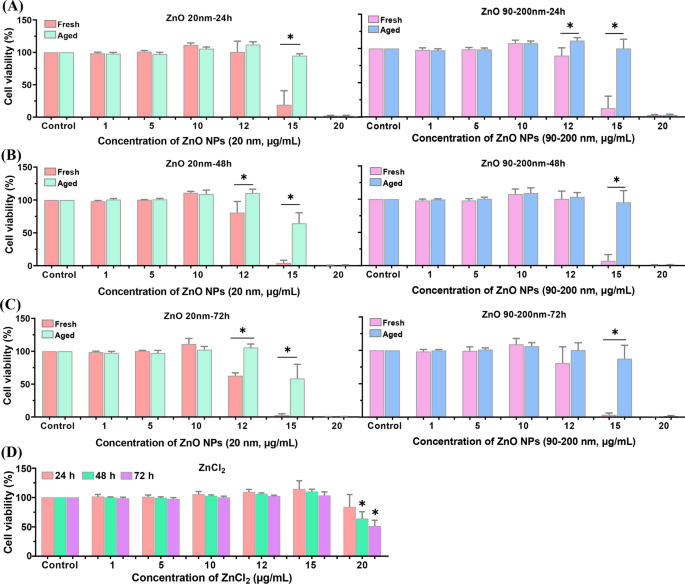
A の新鮮および老化したZnONPによって誘導される細胞生存率 L 細胞。 A L 細胞をさまざまな濃度の新鮮なおよび老化したZnONP(20および90〜200 nm)とともに24時間インキュベートしました( A )、48時間( B )および72時間( C )。 D A L 細胞はさまざまな濃度のZnCl 2 にさらされました さまざまな時間に。データは≥3回の独立した実験に基づいており、平均±SD、* p として表されています。 <0.05
新鮮なZnONPの治療はアポトーシス経路を活性化し、アポトーシス遺伝子の発現をアップレギュレーションしました
老化したNPの細胞毒性の低下につながる根本的なメカニズムを明らかにするために、新鮮なZnONPと老化したZnONPの両方からのRNAシーケンスデータを分析しました。図4A、Bに示すように、新鮮なZnO NPで処理した後、アポトーシス経路はJurkat細胞で活性化されました( p =0.017)およびHMDMセル( p =0.041)。アポトーシス遺伝子: ANXA1 、 CYLD 、 TNFSF10 、 IER3 、 CDKN1A 、 6月 、 SAT1 、 PMAIP1 、 CD38 および ISG20 新鮮なZnONPで処理されたJurkat細胞が大幅に濃縮されました。アポトーシス遺伝子: CD38 、 TNFRSF12A 、 CCNA1 、 BMP2 、 PPP2R5B 、 EREG 、 IFNGR1 、 CD44 、 CD14 、 GNA15 、 GCH1 、 TIMP1 、 BTG2 、 IL1B 、 IL1A 、 BTG3 、 BCL2L11 、 SC5D および SPTAN1 新鮮なZnONP処理HMDM細胞で有意に濃縮されました(図4C、D)。ジャーケット細胞(末梢血Tリンパ球細胞)とHMDM細胞(ヒト単球由来マクロファージ)は異なる種類の細胞であるため、アポトーシスを引き起こす方法は異なる可能性があります。要約すると、これらの結果は、新鮮なZnO NPの曝露が、さまざまな種類の細胞でさまざまなアポトーシス経路を活性化する可能性があることを示しています。
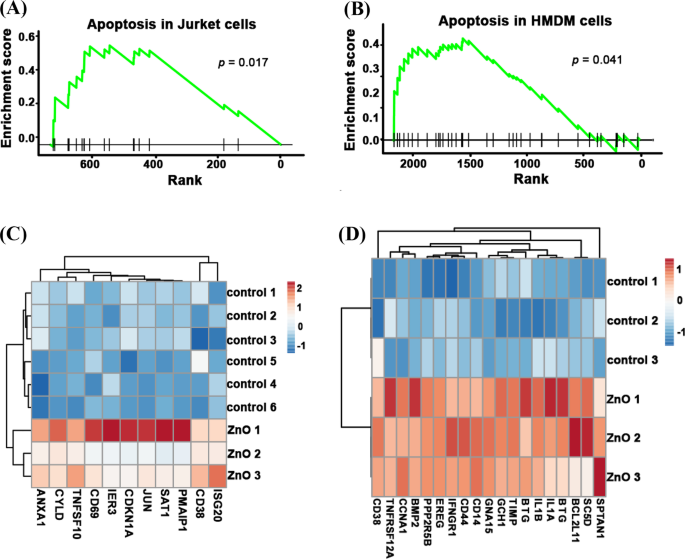
アポトーシス経路は、新鮮なZnONP処理ジャーケットおよびHMDM細胞のRNA-seqデータで強化されました。新鮮なZnONP処理ジャーケット細胞のアポトーシス経路から有意に発現した遺伝子の濃縮スコア( A )およびHMDMセル( B )。新鮮なZnONP処理ジャーケット細胞のアポトーシス遺伝子発現のヒートマップ( C )およびHMDMセル( D )とその対照群
老化したZnONPはアポトーシス遺伝子の発現を新鮮なZnONPとしてアップレギュレートしませんでした
老化したZnONP処理された A からのRNAシーケンスデータ L 細胞は、p53、PI3k–Akt、FoXO、グルタチオン、ErbB、HIF-1、オキシトシン、およびJak-STATシグナル伝達経路が濃縮されていることを示しました(図5A)。 JurketおよびHMDM細胞で濃縮されたアポトーシス遺伝子は、老化したZnO NP処理細胞では有意な影響を受けませんでした(図5B)。調査結果をさらに確認するために、リアルタイムPCRによって関連遺伝子の発現をテストしました。アポトーシス遺伝子のいくつかが見つかりました: BMP2 、 PMAIP1 、IL1α 、 CD69 、 CCNA1 、 CD38 およびIL1β 老化したZnONPで処理された A では検出できませんでした L おそらくこれらの遺伝子のほとんどが免疫系細胞で発現しているためです。その他のアップレギュレーションされたアポトーシス遺伝子(IL1α 、IL1β および CD59 )新鮮なZnO NP処理群で観察されたものは、老化したZnONP処理によって発現レベルに有意な変化はありませんでした。 MT1 ポジティブコントロールとして機能するものは、発現レベル、カスパーゼ3 の発現が大幅に増加しました。 大幅な変更はありませんでした(図5C)。これらのデータは、老化したZnO NPは、それらの新鮮な対応物とは異なり、 A のアポトーシス経路遺伝子を活性化する効果が低いことを示唆しています。 L セル。
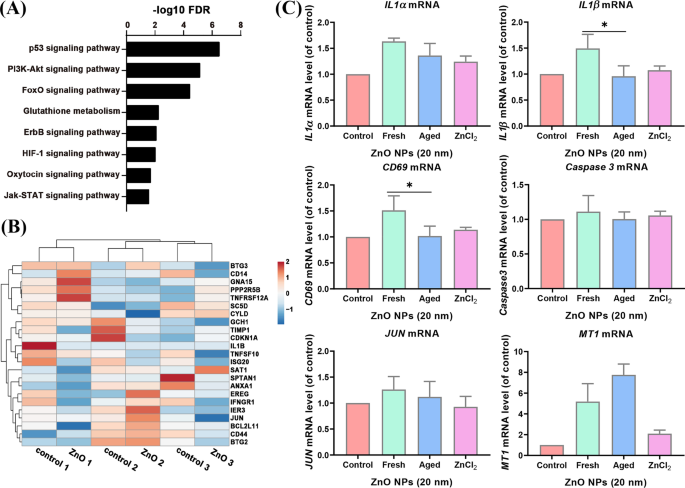
アポトーシス経路は、老化したZnONP処理された A のRNA-seqデータでは濃縮されませんでした。 L 細胞。 ( A )老化したZnONP処理された A からの濃縮経路の遺伝子オントロジー分析 L 細胞。 ( B )老化したZnONP処理 A のアポトーシス遺伝子発現のヒートマップ L 細胞と対照群。 ( C )選択されたアポトーシス遺伝子とコントロール遺伝子の発現( MT1 )新鮮で熟成したZnONP処理 A L セル
新鮮であるが老化していないZnONPは、活性化カスパーゼ3タンパク質の発現レベルを増加させました
カスパーゼ3 の遺伝子発現の検出 単独では、アポトーシス経路の活性化を直接示すことはできません。 ZnO NPの処理によってアポトーシスタンパク質のレベルが変化するかどうかをさらに分析するために、細胞アポトーシスの活性化を示すために一般的に使用されるバイオマーカーである切断型カスパーゼ3タンパク質の発現をウエスタンブロッティング分析で調べました。図6に示すように、対照群と比較して、新鮮なZnO NP(20 nm)処理により、切断されたカスパーゼ3タンパク質の細胞レベルが1.31±0.023倍増加しました。これは、老化した20 nm ZnONPよりも有意に高かった-治療群(1.12±0.039倍)。新鮮なZnONPの粒子サイズを90〜200 nmに増やすと、新鮮なNPによって誘導される切断されたカスパーゼ3タンパク質の発現は、1.46±0.078倍に増加し、老化したNPの発現よりも大幅に大きくなりました(1.07±0.075倍)。 。これらのデータは、老化した対応物と比較して、細胞アポトーシスの誘導における新鮮なZnONPのより高い効力をさらに示しています。
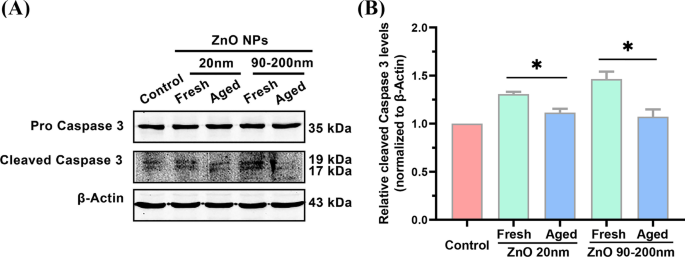
A のアポトーシスレベル L 新鮮で老化したZnONP(20および90〜200 nm)とインキュベートした細胞。ウエスタンブロッティング分析( A )および定量化( B )細胞を12μg/ mLの新鮮および老化したZnONP(20および90〜200 nm)と72時間インキュベートした場合の、切断されたカスパーゼ3タンパク質レベル。データは≥3回の独立した実験に基づいており、平均±SD、* p として表されています。 <0.05
ディスカッション
ZnO NPは、物理化学的変換を受けてZn 5 になることが報告されています。 (CO 3 ) 2 (OH) 6 Zn 2+ の放出を伴う 自然老化の過程で[23、28]。ただし、変換された(老化した)ZnONPによって誘導される細胞毒性とその根底にあるメカニズムは不明なままです。ここでは、新鮮なZnO NPと老化したZnONPの間の多様な細胞毒性のメカニズムを明らかにするために、RNAシーケンス分析とRT-PCRテストを実施しました。また、ウエスタンブロッティングを適用して、細胞アポトーシスの主要な実行因子であるカスパーゼ3のタンパク質レベルを調べました。
私たちのデータは、老化したZnONPが A の新鮮なZnONPよりもはるかに少ない細胞毒性を誘発することを示しました L 細胞。 LC 100 今回の研究では、両方の新鮮なZnO NP(90〜200nmと20nm)の濃度が15μg/ mL未満でした(図3)。これは、LC 100 NIH-3T3またはMSTOセルに対する19〜36nmのZnONPの比率は約15μg/ mLです[29]。 NPの物理化学的特性の環境変化が、NPの毒性を劇的に変化させる可能性があることを確認しました。 ZnO NPの硫化プロセスは、それらの電荷、疎水性、および凝集状態を変化させ、その結果、ヒト唾液、汗および気管支肺胞洗浄液中の硫化物状態NPの吸着をもたらすことが報告されている。そして、ZnO NPによって吸着されたタンパク質は、その生物学的効果に影響を与える特別なタンパク質クラウンを形成しました[19]。生理的溶液(唾液など)に広く存在するリン酸塩は、ZnONPを準安定ZnHPO 4 に変換する可能性があります。 およびZn 3 (PO 4 ) 2 約5〜10時間以内に、消化管上皮細胞に対して細胞毒性を示しました[20]。 Ivask etal。シンクロトロン放射X線吸収を使用して、ヒトTリンパ球(37°C、10%FBSを含む細胞培養培地RPMI1640)へのin vitro曝露システムでのZnO NP(≤3μg/ mL)の完全な変換の発生を証明しました近吸収端構造分光法(XANES)。形質転換産物のスペクトルと細胞毒性は、ZnSO 4 のものと一致していました。 [21]。私たちの結果は、ZnCl 2 の用量依存性および時間依存性の毒性を明らかにしました。 A へ L 細胞毒性は、新鮮なZnO NPと老化したZnO NPの両方よりもはるかに低いです(図3)。この観察結果は、新鮮なZnONPの細胞毒性がその放出されたZn 2+ に完全に起因するわけではないという発見をさらに説明しています。 [30]。
私たちの以前の研究では、老化したZnO NPは、新鮮なZnO NPと比較して、ROS(活性酸素種)を誘発する能力が高く、細胞を殺す能力が弱まっていることも示されました[23]。我々は、老化したZnO NPによって誘発されるより低い細胞毒性が、哺乳動物細胞においてより許容できる可能性があると推論します。 RNAシーケンシングデータの現在の研究は、アポトーシス遺伝子が新鮮なZnO NP処理細胞でアップレギュレーションされており、老化したNP処理グループでは影響がはるかに少ないことを示しています。 IL1αとIL1βはインターロイキン1サイトカインファミリーのメンバーです。 IL1αとIL1βの放出は、カスパーゼ8の部分的に依存するアポトーシスを活性化します[31]。 CD69は、II型膜貫通型受容体のカルシウム依存性レクチンスーパーファミリーのメンバーをコードしています。 CD69発現の増加は、アポトーシスアネキシンVおよびCD95(Fas)マーカーの発現増加と関連していました[32]。 JUNはAP-1転写因子サブユニットです。 JUN活性の増加は、インターロイキン1β変換酵素(ICE)の基質であるα-フォドリンとシステインプロテアーゼのCED-3ファミリーをタンパク質分解的に切断し、プログラム細胞死をさらに引き起こします[33]。これらのアポトーシス遺伝子の発現の増加は、新鮮なNPがいくつかの異なる方法でアポトーシスを引き起こすことを明らかにしました。これらのアポトーシス遺伝子発現の上昇後、アポトーシスプロセスは最終的にアポトーシスタンパク質によって実行されます(図7)。カスパーゼ3は、さまざまなアポトーシスシナリオのコアプロテアーゼです。このタンパク質の切断は、外因性および内因性のアポトーシス経路の両方を活性化するために必要です[34、35]。したがって、切断されたカスパーゼ3の検出は、多種多様なアポトーシスシグナルによって誘導されるアポトーシスを同定するための一般的な方法です[36]。ウエスタンブロッティングデータは、20 nmと90〜200nmの両方のZnONPについて、致死量以下の曝露がすべての治療群でプロカスパーゼ3のレベルを変化させなかったことを明らかにしました。対照的に、切断されたカスパーゼ3は、新鮮なNP処理によって有意に上昇し、老化したNPは、切断されたカスパーゼ3のレベルにほとんど影響を与えませんでした(図6)。 RNA発現解析と組み合わせることで、我々の結果は、細胞アポトーシスの誘導における新鮮なZnONPのより高い効力を明確に解明しました。
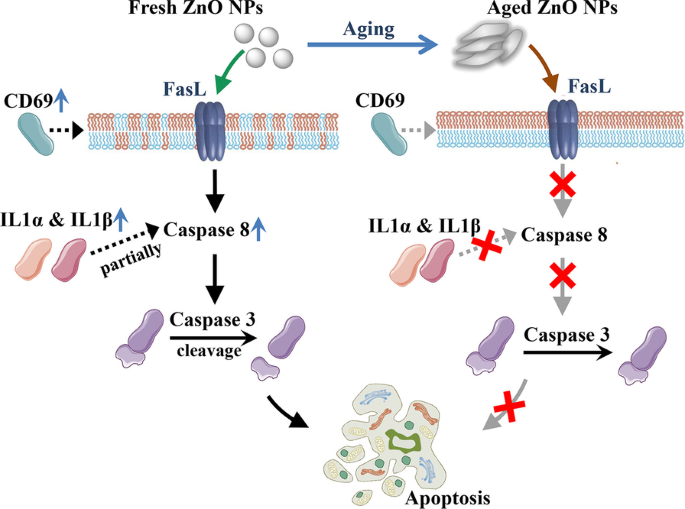
老化したZnONPではなく新鮮なZnONPのモデルは、カスパーゼ8およびカスパーゼ3依存性アポトーシスを誘導します。アポトーシス遺伝子 CD69 の発現増加 新鮮なZnONPに曝露された哺乳動物細胞におけるFasおよびアポトーシスアネキシンVの発現を活性化します。アポトーシス遺伝子IL1αの発現増加 およびIL1β カスパーゼ8依存性アポトーシスを部分的に活性化します。さらに、カスパーゼ3の活性化を引き起こし、アポトーシスを誘導します。 mRNAおよびタンパク質レベルのこれらすべての変化は、老化したZnONPに曝露された哺乳動物細胞では検出できませんでした
結論
本研究では、超純水中でのZnO NPの自然な物理化学的変換を確認し、新鮮なNPと老化したNPによって誘発される細胞毒性の変動を調査しました。老朽化したZnONP処理された A からのRNAシーケンスデータに焦点を当てました L 細胞とデータベースからの新鮮なNPの細胞。新鮮なNPまたはZnCl 2 とは異なる、高齢のNP治療群で特に濃縮されたシグナル伝達経路を比較しました。 治療を受けたグループ。私たちのデータは、老化したZnO NPの細胞毒性の低下は、アポトーシスを誘導する能力の低下と密接に関連している一方で、NPによって活性化される複数の経路の転写調節は、哺乳類細胞における細胞恒常性の確立を促進することを示しています。
データと資料の可用性
該当なし。
略語
- NP:
-
ナノパウダー
- ZnO:
-
酸化亜鉛
- Zn 5 (CO 3 ) 2 (OH) 6 :
-
水亜鉛土
- Zn(OH) 2 :
-
水酸化亜鉛
- ZnCl 2 :
-
塩化亜鉛
- ZnSO 4 :
-
硫化亜鉛
- ZnHPO 4 :
-
リン酸亜鉛亜鉛
- Zn 3 (PO 4 ) 2 :
-
リン酸亜鉛
- A L セル:
-
ヒト-ハムスターハイブリッド細胞
- CHOセル:
-
チャイニーズハムスター卵巣細胞
- ジャーケットセル:
-
末梢血Tリンパ球細胞
- HMDMセル:
-
ヒト単球由来マクロファージ
- NIH-3T3cells:
-
マウス胚性細胞
- MSTOセル:
-
ヒト肺がん細胞
- RPMI1640:
-
ロズウェルパーク記念研究所1640
- ICE:
-
インターロイキン1β変換酵素
- CED-3:
-
線虫Caenorhabditiselegans 死の遺伝子
- IL1α:
-
インターロイキン1alpha
- IL1β:
-
インターロイキン1β
- mRNA:
-
メッセンジャーリボ核酸
- cDNA:
-
相補的なデオキシリボ核酸
- FBS:
-
ウシ胎児血清
- TEM:
-
透過型電子顕微鏡
- XRD:
-
X線回折
- RT-PCR:
-
リアルタイムポリメラーゼ連鎖反応
- CPI:
-
ナノテクノロジー消費者製品在庫
- XANES:
-
放射光X線吸収端構造分光法
- RIPA:
-
ラジオ免疫沈降アッセイ
- SDS-PAGE:
-
ポリアクリルアミドゲル電気泳動
- PVDF:
-
ポリフッ化ビニリデン
- ECL:
-
強化された化学発光
- CCK-8:
-
細胞計数キット-8
ナノマテリアル
- 生体適合性FePO4ナノ粒子:ドラッグデリバリー、RNA安定化、および機能的活性
- Al2O3 / ZnOナノラミネートの形態的、光学的、および電気的特性に及ぼす二重層の厚さの影響
- 界面層の設計によるZnO膜の表面形態と特性の調整
- 銅ナノ粒子の合成と安定化におけるグリーン熟達度:触媒、抗菌、細胞毒性、および抗酸化活性
- ポリオール媒介プロセスによるZnOナノクリップの製造と特性評価
- ラットにおける腹腔内および静脈内投与経路による生合成された銅および酸化亜鉛ナノ粒子の比較invivo精査
- 金属および金属酸化物ナノ粒子のグリーン合成と単細胞藻類Chlamydomonasreinhardtiiに対するそれらの効果
- 酸化亜鉛ナノ粒子の特性と微生物に対するそれらの活性
- 非常に濃縮された亜鉛源を用いた水/グリセロール中のナノサイズの酸化亜鉛の調製のための簡単なアプローチ
- 酸化亜鉛/多孔質陽極酸化アルミナ複合膜の調製とアンチバイオフィルム特性
- インダストリー4.0におけるロボット工学と自動化の役割