


HDAC1を介したMicroRNA-124-5pは、うつ病のラットの学習能力と記憶能力に影響を与えるようにNPYを調節します
要約
うつ病におけるヒストンデアセチラーゼ(HDAC)を中心とした研究は過度に行われていますが、HDAC1についてはあまり行われていません。その中で、本研究は、うつ病におけるHDAC1 / microRNA(miR)-124-5p /ニューロペプチドY(NPY)軸のメカニズムを明らかにするために開始されます。 Sprague Dawleyラットは、慢性的な予測不可能な軽度のストレスによって刺激され、うつ病モデルを確立しました。抑制されたラットに抑制されたHDAC1または抑制されたmiR-124-5pを注射して、体重、学習および記憶能力、酸化ストレスおよび血清中の炎症および海馬組織における神経伝達物質発現におけるそれらの役割を調査した。海馬におけるMiR-124-5p、HDAC1およびNPYの発現を試験した。 miR-124-5p、HDAC1およびNPY発現の相互作用も確認されました。より高いmiR-124-5pとHDAC1およびより低いNPY発現レベルが鬱病ラットの海馬で見られました。抑制されたmiR-124-5pまたは抑制されたHDAC1は、学習能力と記憶能力を弱め、鬱病ラットの体重を増加させました。 miR-124-5pのノックダウンまたはHDAC1の阻害は、酸化ストレスと炎症を抑制し、鬱病ラットの神経伝達物質の発現を促進しました。 HDAC1はmiR-124-5pを介してNPYを制御します。 NPYのノックダウンは、うつ病のラットに対する抑制されたmiR-124-5pの保護効果を無効にしました。私たちの研究は、miR-124-5pまたはHDAC1のいずれかの抑制がNPYをアップレギュレートして、うつ病のマウスの記憶力と学習能力を改善することを示しています。
うつ病とは、世界的に深刻な経済的負担と社会的影響を引き起こす深刻な精神病を無効にすることを指します[1]。うつ病は、患者の全体的な健康を脅かす生理学的、認知的、行動的変化を特徴としています[2]。うつ病に利用できる治療法は、速効性抗うつ薬としてのケタミンの適用と、ニューロンのサブタイプの集団の活動に選択的に対応できる機器の改良によって開発されました[3]。さらに、カーボンドットなどの用途の広いナノアーキテクチャは、神経障害に適用できます[4]。しかし、うつ病の治療が不十分だと、パフォーマンスの低下、行動障害、身体的疾患、薬物乱用、さらには自殺につながる可能性があります[5]。したがって、うつ病の治療における革新的な薬剤を発見することが切実に必要とされています。
microRNA(miR)-124レベルは、うつ病と抗うつ薬を伴う血漿中で上昇することが以前に報告されています[6]。慢性的な予測不可能な軽度のストレス(CUMS)によって誘発されるうつ病の間、海馬でのmiR-124の発現は動的な変化を示し、うつ病のさまざまな段階でのさまざまな病理学的変化を示します[7]。さらに、miR-124は、大うつ病性障害におけるうつ病様行動を改善する候補です[8]。また、miR-124抑制は、マウスのうつ病のような行動の減弱に反映されるように、前頭前野の抗うつ薬として機能します[9]。ヒストンデアセチラーゼ1(HDAC1)は、miR-124 [10]のメディエーターであり、転写調節とエピジェネティック修飾のための多タンパク質複合体を形成します[11]。私たちの知る限り、HDAC1活性の障害は、大うつ病性障害の回復力を促進します[12]。さらに、脳内のHDAC1の抑制は、気分障害やその他の神経可塑性に変化した脳疾患を改善する可能性があります[13]。 HDAC阻害剤は、不安様および飲酒行動を弱める可能性があり、ニューロペプチドY(NPY)の発現を上昇させる可能性があります[14]。また、HDAC活性障害は、扁桃体の中心核および扁桃体の内側核におけるNPY発現の増加につながる可能性があります[15]。 NPYは視床下部の食欲促進性神経ペプチドであり、不安、社会的障害、抑うつ症状を予防するのに十分です[16]。それに加えて、NPYとその受容体は、リポ多糖誘発性うつ病ラットモデルに抗炎症作用と抗うつ作用を及ぼすことが証明されています[17]。まとめると、うつ病におけるHDAC1 / miR-124-5p / NPY軸の相互作用の組み合わせはあいまいです。したがって、この研究は、うつ病治療の治療候補を調査するために、この軸のメカニズムを明らかにすることを目的としています。
この研究に含まれる動物実験は、ハルビン医科大学の第一付属病院の実験動物倫理の要件に準拠していました。実験は、実験動物の摂食条件を改善し、使用する動物の数を減らし、動物の苦痛を軽減するために最適化されました。
特定病原体除去(SPF)グレードのSprague–Dawley(SD)雄ラット、体重200〜220 gは、ハルビン医科大学(中国、ハルビン)の第一付属病院の動物研究センターから提供されました。ラットはSPFグレードの環境(22〜24°C、湿度60〜64%、12時間の明暗サイクル)で飼育されました。モデリング中の期間と他の特定の時点を除いて、ラットは自由に水と餌を手に入れることができました。
モデル化されたラットはランダムに8つのグループに分けられました( n =10):正常グループ(治療なしのラット)、CUMSグループ(CUMS誘発性うつ病ラット)、sh陰性対照(NC)グループ(sh-HDAC1レンチウイルスNCを注射したCUMS誘発性うつ病ラット)、sh-HDAC1グループ(sh-HDAC1レンチウイルスを注射したCUMS誘発性うつ病ラット)、anti-miR-NCグループ(anti-miR-124-5pレンチウイルスNCを注射したCUMS誘発性うつ病ラット)、anti-miR-124-5pグループ(CUMS-抗miR-124-5pレンチウイルスを注射した誘発性うつ病ラット)、anti-miR-124-5p + sh-NCグループ(抗miR-124-5pレンチウイルスおよびsh-NPYレンチウイルスNCを注射したCUMS誘発性うつ病ラット) 、およびanti-miR-124-5p + sh-NPYグループ(抗miR-124-5pレンチウイルスおよびsh-NPYレンチウイルスを注射したCUMS誘発性うつ病ラット)。
CUMS誘発性うつ病ラットモデルは、ウィルナーの方法によって確立されました[18]。正常群のラットは刺激なしで餌と水を与えられたが、他の7群のラットは独立したケージで35日間のCUMSを受けた。これらのラットは、コンテナ(水深30cm)内で4°Cで5分間氷水で泳ぐこと、昼と夜の反転(日中は暗闇に12時間、明るい空間に照らされたラット)によって刺激されました。白熱灯(夜間12時間)、ツバメテールクリップによる30秒間のテールクランプ、1分間の振とう、24時間の絶食と水分欠乏、30分間の超音波刺激(50 Hz)、ウェットパッド、および空調17℃で。ラットはこれらの刺激の1つによって毎日ランダムに刺激され、同じ刺激が合計で5回以下適用されました。
ラットを2%ペントバルビタールナトリウム(50 mg / kg)で麻酔し、両側海馬(前後径=4.8 mm、中外側距離=±2.5 mm、後腔と大泉門の間の距離=− 3.5 mm)にレンチウイルスを1で注射しました。 26Gニードルを備えたハミルトンマイクロシリンジによるμL/分。逆流を防ぐために、針を注射部位に5分間保持した。頭蓋骨を骨ワックスで密封し、切開部を縫合し、ラットを7日間回復させた[19]。 sh-HDAC1レンチウイルスとそのNC、anti-miR-124-5pレンチウイルスとそのNC、およびsh-NPYとそのNC(力価:10
8
TU / mL)は、GenePharma Co. Ltd.(上海、中国)から購入しました。
ラットは、CUMSモデリングの前日(0日)、CUMSモデリング後(36日目)、およびレンチウイルス干渉後(52日目)に体重を測定しました。
オープンフィールド試験(OFT)は、抑うつ行動を評価するために一般的に使用されていた新しい環境でのラットの自律神経および探索行動を検出できました。このテストは、ラットの活動を記録するためにオープンフィールドボックスの上に配置されたビデオ追跡システムを備えた黒い木製のオープンフィールドボックス(100cm×100cm×50cm)で実施されました。開いた箱の底は白い線で25グリッド(20cm×20cm)に分割されました。ラットを事前に定義されたランダムな順序で中央グリッドに配置し、グリッドを横切る量(ラットが四肢または片方の後肢を備えた二重前肢でグリッドに入る)および飼育の発生率(ラットが前肢を持ち上げ、1つで直立する)後肢)は、ビデオ追跡システムによって記録されました。このテストは、静かで薄暗い屋内環境で実行されました。すべてのテストの後、臭いを取り除くためにオープンフィールドボックスを75%アルコールで洗浄しました。
無快感症は、うつ病の重大な症状の1つでした。ショ糖嗜好性試験(SPT)は、ラットの抑うつ様行動を評価することができます。 SPTの前に、各ラットに2本の砂糖水(1%スクロース、w / w)を24時間提供し、1本の滅菌水と1本の砂糖水をさらに24時間提供しました。その後、ラットのショ糖嗜好性パーセンテージ(SPP)を次のように検出しました:23時間の絶食と水分欠乏の後、ラットに1本の滅菌水と1本の砂糖水(1%ショ糖)を30分間与えました。 1時間後、2本のボトルの位置を逆にし、ラットはこれらの2本のボトルをさらに30分間自由にした。この1時間の間に、これら2本の滅菌水と砂糖水(1%スクロース)の消費量(mL)が測定され、SPPが計算されました。 SPP =砂糖水消費量/(砂糖水消費量+滅菌水消費量)×100%。
モリス水迷路試験(MWM)は、ラットの空間学習と記憶能力を評価するために広く適用されました。 MWM試験は、水深40cmの円形水槽(直径150cm、高さ60cm)で実施しました。水温は22±1°Cに維持され、水は無毒でお手入れが簡単な染料で黒く染められました。タンクと水面は4つの象限(SE、SW、NW、NEの象限)に分割され、各象限は対応するアイコンで明るい色と特別な形で内壁に装飾されています。ターゲットプラットフォームは、NE象限の中心に位置し、水面下1.5 cmに浸された、円形の透明なプラットフォーム(直径11 cm)でした。ビデオ追跡システムがタンクの中央の上に設置され、水泳の速度、経路、およびラットがターゲットプラットフォームに入り、象限で泳ぐのに費やした時間を記録しました。ラットはランダムな順序で4つの象限から水中に置かれ、60秒以内にターゲットプラットフォームを探索して搭乗することができました。ラットがプラットフォームに乗るのにかかった時間は、逃避潜時として記録された。ラットが60秒以内にターゲットプラットフォームに搭乗できなかった場合、ラットは誘導され、逃避潜時は60秒として記録されました。各試験後、ラットを標的プラットフォーム上に15秒間留まらせ、ラットを1日4回5日間訓練した。最終的な脱出潜時は、3日から5日の脱出潜時の平均でした。 MWM試験の6日目に、宇宙探査試験が実施されました。ターゲットプラットフォームは水タンクから取り外され、ラットは南西象限から水に入った。 60秒以内に北東象限で泳ぐラットの時間を宇宙探査時間として記録しました。
この実験のタイムラインを図1に示します。
本研究の年表
最後のMWM試験の1日後、ラットを安楽死させ、2%ペントバルビタールナトリウム(50mg / kg)を腹腔内注射した。胸腔を開いて注射器で心臓の血液を採取し、3500 r / minで15分間3回遠心分離し、上清を-20°Cで保存しました。採血後、左心室から大動脈にカテーテルを挿入し、500mLの生理食塩水を灌流して全血を流しました。海馬組織を分離し、1.5 mLの遠心分離管に入れ、秤量し、-80°Cで保存しました。海馬の一部を4%パラホルムアルデヒドで2時間固定し、グラジエントスクロースで脱水し、パラフィンに包埋しました。
スーパーオキシドジスムターゼ(SOD)、マロンジアルケヒチェ(MDA)、グルタチオン(GSH)、インターロイキン(IL)-β、腫瘍壊死因子-α(TNF-α)、および一酸化窒素(NO)濃度を検出するために、血清を再加温しました。 SOD、MDA、およびGSHを検出するためのキットは、Beyotime Institute of Biotechnology(上海、中国)から提供され、IL-β、TNF-α、およびNOを検出するための酵素結合免疫吸着測定法(ELISA)キットは、NanJing JianCheng Bioengineering Institute(南京、中国)。
パラフィン切片を脱ロウし、エタノールで水和し、蒸留水ですすぎ、ヘマトキシリン染色液で20分間染色した。その後、切片が青色になるまで水道水ですすいだ。次に、切片を1%エタノール性塩酸溶液に10秒間入れ、水道水ですすぎ、エタノールで脱水した後、エオシンで2分間染色し、高濃度アルコールで脱水し、キシレンで透過処理した。最後に、切片を密封し、生物学的顕微鏡で観察しました。
海馬組織の重さを量り、通常の生理食塩水(100μL/ 10mg)中の超音波によって均質化した。ホモジネートを4°Cで30分間保持し、12,000 rpm(4°C)で3分間遠心分離して、上清を回収しました。上清のタンパク質濃度はビシンコニン酸(BCA)タンパク質検出キット(CWBIO、北京、中国)で検出され、ノルエピネフリン(NE)、セロトニン(5-HT)、ドーパミン(DA)の発現レベルはELISAキットで検出されました。 NE、5-HT、およびDA ELISAキットは、Liweiping Biotechnology Co.、Ltd。(北京、中国)から提供されました。
RNAは、RNA抽出キット(Promega、マディソン、ウィスコンシン州、米国)によって海馬組織から抽出され、RNAの濃度と純度は紫外分光光度計によって検出されました。逆転写キットの説明書(DRR047S、タカラ、大連、中国)に続いて、相補DNA(cDNA)へのRNA逆転写を行いました。 mRNAはGoldScriptワンステップRT-PCRキット(Applied Biosystems、Carlsbad、CA、USA)によってcDNAに逆転写され、miRNAはHairpin-it™miRNA定量検出キット(GenePharma)によって逆転写されました。 RT-qPCR検出キット(Promega)を適用して、組織内のHDAC1、miR-124-5p、およびNPYの発現を検出しました。 U6はmiR-124-5pの内部対照として示され、β-アクチンはHDAC1およびNPYの内部対照として示されました。 PCRプライマーは、Sangon Biotech Co.、Ltd。(Shanghai、China)から入手しました(表1)。標的遺伝子の相対的発現は2
-△△
によって計算されました Ct法。
海馬組織を氷上で細かく切断し、放射免疫沈降アッセイ溶解物(北京ソーラーバイオサイエンスアンドテクノロジー株式会社、北京、中国)によって溶解してタンパク質を抽出した。タンパク質濃度はBCA法により測定した。全タンパク質(50μg)に5×ドデシル硫酸ナトリウム(SDS)ローディングバッファーを1:4で加え、沸騰水浴で5分間加熱しました。次に、タンパク質をドデシル硫酸ナトリウムポリアクリルアミドゲル電気泳動にかけ、膜にエレクトロブロッティングし、5%ミルクで1時間ブロックしました。その後、タンパク質をHDAC1(1:1000、Cell Signaling Technology、マサチューセッツ州ビバリー、米国)、NPY(1:800、NeoMakers、米国カリフォルニア州フリーモント)、およびβ-アクチン(1: 1000、Abcam、Cambridge、MA、UK)一晩、西洋ワサビペルオキシダーゼ標識二次抗体で再プローブしました。メンブレンは強化された化学発光によって開発され、光学密度はQuantityOneグレー分析ソフトウェアによって計算されました。標的遺伝子のタンパク質発現は、β-アクチングレー値に対するグレー値の比率として表されました。
ChIPアッセイは、EZ-ChIPキット(Millipore、米国マサチューセッツ州ベッドフォード)の指示に従って実施しました。 HEK293T細胞を1%ホルムアルデヒドとともに10分間インキュベートし、グリシンで終結させました。次に、細胞を2000rpmで5分間遠心分離し、超音波処理のためにSDS溶解緩衝液を加えた。 10,000 g、4℃で10分間遠心分離し、細胞(100μL)を900μLのChIP希釈バッファー、20μLの50×PICおよび60μLのProteinAアガロース/サーモン精子DNAと4℃で1時間反応させました。 10分間放置しました。沈殿物を700rpmで1分間遠心分離し、20μLを投入しました。チューブにHDAC1抗体(1μL)と免疫グロブリンG抗体を加え、もう一方のチューブに抗体を加えませんでした。 2本のチューブ内のサンプルを一晩インキュベートし、溶出し、脱架橋しました。 DNAを取得した後、サンプルをRT-qPCRでテストしました。
バイオインフォマティクスのウェブサイト(https://cm.jefferson.edu/rna22/Precomputed/)は、miR-124-5pとNPYの結合部位を分析しました。 NPY-野生型(WT)およびNPY-変異体(MUT)プラスミドは、Huayueyang Biotechnology Co.、Ltd。(北京、中国)によって生成されました。 mimicNCまたはmiR-124-5pmimicと組み合わせて、プラスミドをHEK293T細胞にトランスフェクトしました。細胞ルシフェラーゼ活性は、ルシフェラーゼ検出キット(BioVision)およびGlomax20 / 20ルミノメーター(Promega)によって測定されました。
データ分析には、SPSS 21.0統計ソフトウェア(IBM Corp. Armonk、NY、USA)を使用しました。結果は平均±標準偏差として表されました。 2つのグループ間の比較は、 t によってテストされました。 テスト。複数のグループ間の比較は、一元配置分散分析(ANOVA)によって評価され、その後、テューキーの事後検定によるペアワイズ比較が行われました。 P 両面テストと P を表しました <0.05は統計的有意性があると見なされました。
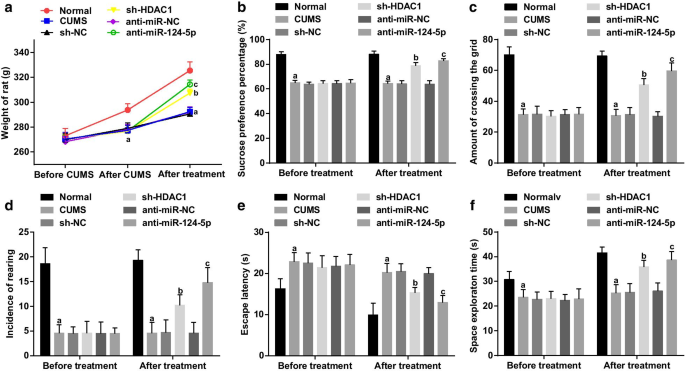
ラットは、CUMSモデリングの1日前(0日目)、モデリングの停止後1日目(36日目)、およびレンチウイルス干渉の7日後(52日目)に体重を測定しました。モデル化前の各グループのラットの体重に差は検出されませんでした(すべての P > 0.05)。モデリング後、ラットの体重は、CUMSグループ、sh-NCグループ、sh-HDAC1グループ、anti-miR-NCグループ、およびanti-miR-124-5pグループで、正常グループと比較して減少しました(すべて P > <0.05)。 CUMS群、sh-NC群、sh-HDAC1群、anti-miR-NC群、anti-miR-124-5p群(すべて P )のラット体重に差異は認められなかった。> 0.05)。干渉後、ラットの体重は、正常群と比較してCUMS群で減少しました(すべて P <0.05)。 CUMSグループ、sh-NCグループ、およびanti-miR-NCグループ(すべて P )で、ラットの体重に差異は見られませんでした。> 0.05)。ラットの体重は、sh-HDAC1グループとanti-miR-124-5pグループのNCグループ(両方 P )と比較して増加しました。 <0.05)、HDAC1またはmiR-124-5pの阻害により、うつ病のラットの体重が増加したことを示しています(図2a)。
HDAC1またはmiR-124-5pのいずれかを阻害すると、体重が増加し、うつ病のラットの学習能力と記憶能力が向上します。 a うつ病ラットの体重に対するHDAC1またはmiR-124-5pによる阻害の影響; b HDAC1またはmiR-124-5pによる阻害前後のSPP; c HDAC1またはmiR-124-5pによる抑制の前後にOFTでグリッドを通過する頻度。 d HDAC1またはmiR-124-5pによる阻害の前後のOFTでの飼育の発生率; e HDAC1またはmiR-124-5pによる抑制の前後のMWMテストでの待ち時間を回避します。 f HDAC1またはmiR-124-5pによる抑制前後のMWMテストでの宇宙探査時間。 n =10; P 正常群と比較して<0.05; b P sh-NCグループと比較して<0.05; c P 抗miR-NCグループと比較して<0.05。複数のグループ間の比較は一元配置分散分析によって評価され、ペアワイズ比較はテューキーの事後検定によって評価されました
SPT、OFT、MWMテストを適用して、SPP、グリッドを通過する頻度、飼育の発生率、および脱出潜時と宇宙探査時間を検出しました。干渉前(図2b–f)は、通常のグループと比較して、SPP、グリッドを通過する頻度、飼育の発生率、および宇宙探査時間が減少したのに対し、CUMSグループのsh-NCでは逃避潜時が延長したことが概説されました。グループ、sh-HDAC1グループ、anti-miR-NCグループ、anti-miR-124-5pグループ(すべて P <0.05)、ラットにおけるうつ病のような行動の発達を示唆している。 CUMSグループ、sh-NCグループ、sh-HDAC1グループ、anti-miR-NCグループ、およびanti-miR-124の間で、SPP、グリッドを通過する頻度、飼育の発生率、宇宙探査時間、および脱出潜時に差異はありませんでした。 -5pグループ(すべて P > 0.05)。干渉後、sh-NCグループおよびanti-miR-NCグループと比較して、SPP、グリッドを通過する頻度、飼育の発生率および宇宙探査時間は増加しましたが、sh-HDAC1グループおよびanti-miR-124-では脱出潜時が短縮されました。 5pグループ(すべて P <0.05)。 CUMSグループ、sh-NCグループ、およびanti-miR-NCグループ(すべて P )の間で、SPP、グリッドを通過する頻度、飼育の発生率、宇宙探査時間、および脱出潜時に差は観察されませんでした。> 0.05)、HDAC1のサイレンシングまたはmiR-124-5pの抑制が、うつ病のような行動を弱め、ラットの学習能力と記憶能力を改善する可能性があることを示唆しています。
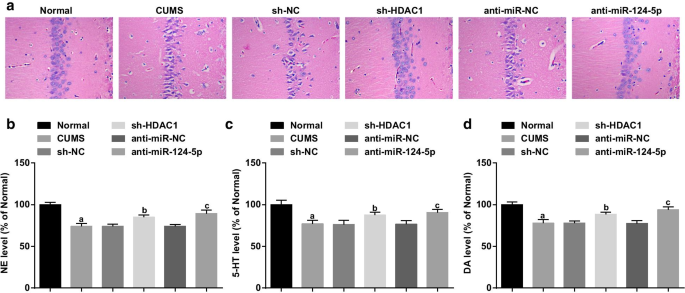
HE染色による海馬病変の観察(図3a)は、正常群のラットの海馬にきちんと配置されたニューロンが、明確な形態、正常な構造、密な層、明確な細胞核、および明らかな核小体を示したことを明らかにした。 CUMSグループのラットのニューロンは収縮し、数が減少し、クロマチンが不均一に分布し、層が薄くなってゆるく配置されていました。 sh-NCおよびanti-miR-NCグループのラットは、CUMSグループと同じ状況を示しました。 sh-HDAC1およびanti-miR-124-5pグループのラットは、NCグループと比較して、ニューロンの順序が増加し、損傷が軽減されました。 HDAC1のノックダウンまたはmiR-124-5pの阻害により、うつ病ラットの海馬病変が軽減されることが示されました。
HDAC1またはmiR-124-5pのいずれかの阻害は、病的な海馬の損傷を軽減し、うつ病のラットにおける神経伝達物質の発現をアップレギュレートします。 a うつ病ラットの海馬病変のHE染色; b 海馬組織におけるNE発現のELISA; c 海馬組織における5-HT発現のELISA; d 海馬組織におけるDA発現のELISA; n =6; P 正常群と比較して<0.05; b P sh-NCグループと比較して<0.05; c P 抗miR-NCグループと比較して<0.05。複数のグループ間の比較は一元配置分散分析によって評価され、ペアワイズ比較はテューキーの事後検定によって評価されました
うつ病は、脳の神経伝達物質障害に関連していた。したがって、ラット海馬のDA、NE、および5-HT神経伝達物質のレベルをELISAで測定しました。結果は、対照的に、正常群では(図3b–d)、DA、NE、および5-HTのレベルの低下が、CUMS群、sh-NC群、sh-HDAC1群、anti-miRのラットで見られたことを示しました。 -NCグループおよびanti-miR-124-5pグループ(すべて P <0.05)。 CUMSグループ、sh-NCグループ、anti-miR-NCグループ(すべて P )の神経伝達物質の発現に違いは見られませんでした。> 0.05)。 sh-NCおよびanti-miR-NCグループと比較して、DA、NE、および5-HTレベルは、sh-HDAC1およびanti-miR-124-5pグループ(すべて P )のラットで増加しました。 <0.05)、HDAC1のダウンレギュレーションまたはmiR-124-5pの減少が、うつ病ラットの海馬におけるDA、NE、および5-HTのレベルをアップレギュレーションする可能性があることを意味します。
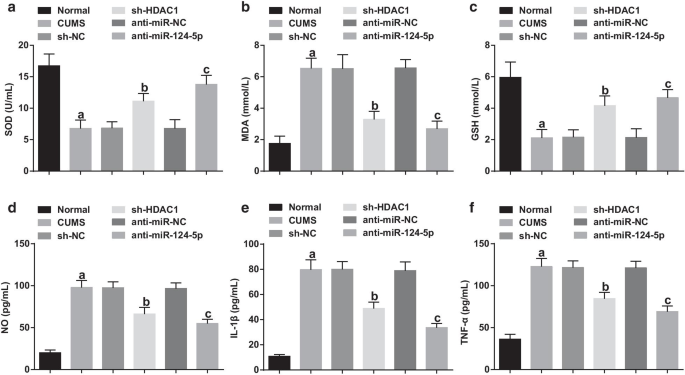
血清中の酸化ストレス関連および炎症性因子の発現を測定した。正常群に関しては、SODおよびGSH活性が損なわれ、MDA、IL-1β、TNF-α、およびNOレベルは他のうつ病モデル群(すべて P )で増加しました。 <0.05)。 SODおよびGSH活性、ならびにCUMSグループ、sh-NCグループおよびanti-miR-NCグループ間のMDA、IL-1β、TNF-αおよびNOのレベルは、差を示さなかった(すべての P > 0.05)。 sh-NC群およびanti-miR-NC群と比較して、SODおよびGSH活性の増加が認められ、sh-のMDA、IL-1β、TNF-α、およびNOレベルの減少が観察されました。 HDAC1グループおよびanti-miR-124-5pグループ(すべて P <0.05)(図4a–f)、HDAC1の枯渇またはmiR-124-5pのダウンレギュレーションが、うつ病ラットの酸化ストレスと炎症を軽減できることを示しています。
HDAC1またはmiR-124-5pのいずれかを阻害すると、うつ病のラットの酸化ストレスと炎症が抑制されます。 a – c うつ病ラットの血清中のSOD、MDAおよびGSH濃度に対するHDAC1またはmiR-124-5pの阻害の影響。 d – f うつ病ラットの血清中のIL-1β、TNF-α、およびNO濃度に対するHDAC1またはmiR-124-5pの阻害の影響。 n =10; P 正常群と比較して<0.05; b P sh-NCグループと比較して<0.05; c P 抗miR-NCグループと比較して<0.05。複数のグループ間の比較は一元配置分散分析によって評価され、ペアワイズ比較はテューキーの事後検定によって評価されました
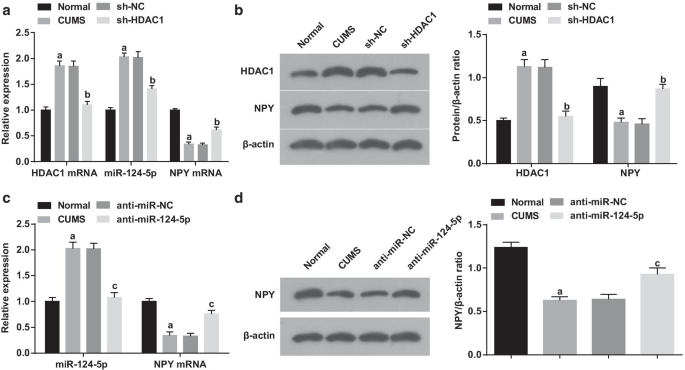
海馬におけるHDAC1、miR-124-5pおよびNPYの検出には、RT-qPCRおよびウエスタンブロット分析を採用しました。 CUMSグループ(すべて P )では、正常グループと比較して(図5a、b)、HDAC1とmiR-124-5pが増加し、NPYが減少したことが概説されました。 <0.05)。 HDAC1、miR-124-5p、およびNPYの発現は、CUMSグループとsh-NCグループ(すべて P )で変化を示しませんでした。> 0.05)。 sh-NCグループと比較して、sh-HDAC1グループは、HDAC1とmiR-124-5pの減少、およびNPY発現レベルの上昇(すべて P )に反映されていました。 <0.05)。上記の調査結果は、レンチウイルス干渉の成功と、miR-124-5pとHDAC1発現の間の正の関係を示唆しています。
HDAC1はmiR-124-5pを仲介してNPYを制御します。 a RT-qPCRによるHDAC1の阻害後の海馬組織におけるHDAC1、miR-124-5pおよびNPY mRNAの発現( n =6); b ウエスタンブロット分析によるHDAC1の阻害後の海馬組織におけるHDAC1およびNPYタンパク質の発現( n =6); c RT-qPCRによるmiR-124-5pの阻害後の海馬組織におけるMiR-124-5pおよびNPYmRNAの発現( n =6); d ウエスタンブロット分析によるmiR-124-5pの阻害後の海馬組織におけるNPYタンパク質発現( n =6); P 正常群と比較して<0.05; c P sh-NCグループと比較して<0.05。複数のグループ間の比較は一元配置分散分析によって評価され、ペアワイズ比較はテューキーの事後検定によって評価されました
また、miR-124-5p抑制後のmiR-124-5pおよびNPYの発現が検出され、結果(図5c、d)は、正常群と比較して、miR-124-5pの上昇およびNPYの低下がCUMSグループ(両方の P <0.05)。それどころか、anti-miR-124-5pグループは、anti-miR-NCグループ(両方 P )と比較して、miR-124-5pが減少し、NPYが上昇する傾向がありました。 <0.05)。 CUMSグループとanti-miR-NCグループ(両方 P )のmiR-124-5pとNPYの発現に矛盾は認められませんでした。> 0.05)。結果は、レンチウイルス干渉の成功と、NPYとmiR-124-5pの間の負の関係を示していました。
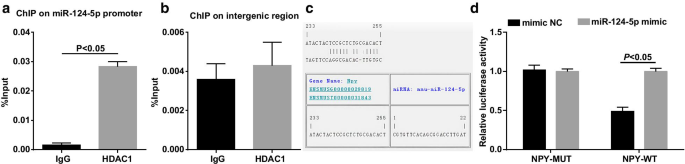
ChIPアッセイは、HDAC1がmiR-124-5pプロモーターに結合できるかどうかをテストすることであり、結果は(図6a、b)HDAD1がmiR-124-5pプロモーター( P > <0.001)、HDAC1がmiR-124-5pを直接制御できることを示します。
HDAC1はmiR-124-5pのプロモーターに結合します。 a ChIPアッセイによるHEK293T細胞のHDAC1およびmiR-124-5pプロモーター領域の結合量( n =3); b ChIPアッセイによるHEK293T細胞のHDAC1と無関係の遺伝子間領域の結合量( n =3); c 生物学情報ソフトウェアのウェブサイトによるMiR-124-5pおよびNPY結合部位。 d デュアルルシフェラーゼレポーター遺伝子アッセイによるmiR-124-5pとNPYのターゲティング関係( n =3); 2つのグループ間の比較は、 t によってテストされました。 テスト
miR-124-5pとNPYの間のターゲティング関係は、RNA22ツールとデュアルルシフェラーゼレポーター遺伝子アッセイによって予測および検証されました(図6c、d)。 With respect to the cells co-transfection with NPY-3′UTR-WT and mimic NC, the cells with co-transfection of NPY-3′UTR-WT and miR-124-5p mimic showed impaired luciferase activity (P <0.05)。 No difference was recognized in the luciferase activity in the cells co-transfected with NPY-3′UTR-MUT and mimic NC, and the cells co-transfected with NPY-3′UTR-MUT and miR-124-5p mimic (P > 0.05).
Spontaneous depletion of NPY and miR-124-5p was programmed to explore their interplay in depressed rats. It was exhibited that (Fig. 7a, b) lower NPY expression level was noticed in the anti-miR-124-5p + sh-NPY group versus the anti-miR-124-5p + sh-NC group (P <0.05)。 Body weight and behavioral function tests illustrated that (Fig. 7c–f) by comparison with the anti-miR-124-5p + sh-NC group, the rats in the anti-miR-124-5p + sh-NPY group presented reduced weight, SPP, frequency of crossing the grid, incidence of rearing and space exploration time with increased escape latency (all P <0.05)。 HE staining of the hippocampal lesions pictured that (Fig. 8a) in comparison with the anti-miR-124-5p + sh-NC group, the number of hippocampal neurons was reduced, and hippocampal neurons were darkly stained, sparsely and disorderly arranged in an irregular shape with reduced cell layers in the anti-miR-124-5p + sh-NPY group. Moreover, versus the anti-miR-124-5p + sh-NC group, the rats in the anti-miR-124-5p + sh-NPY group was accompanied by reduced DA, NE and 5-HT (all P < 0.05) (Fig. 8b). Besides, impaired SOD and GSH activities, and increased MDA, IL-1β, TNF-α and NO levels existed in the rats of the anti-miR-124-5p + sh-NPY group versus the anti-miR-124-5p + sh-NC group (all P < 0.05) (Fig. 8c, d). Collectively, knockdown of NPY abolished the protective effects of repressed miR-124-5p on depressed rat.
Knockdown of NPY abolishes the effects of inhibited miR-124-5p on body weight and behavioral function of depressed rats. a MiR-124-5p and NPY mRNA expression in hippocampal tissues after inhibition of both miR-124-5p and NPY by RT-qPCR (n = 6); b NPY protein expression in hippocampal tissues after inhibition of both miR-124-5p and NPY by western blot analysis (n = 6); c Body weight of depressed rats after inhibition of both miR-124-5p and NPY (n = 10); d SPP of depressed rats after inhibition of both miR-124-5p and NPY (n = 10); e Frequency of crossing the grid and incidence of rearing after inhibition of both miR-124-5p and NPY (n = 10); f Escape latency and space exploration time after inhibition of both miR-124-5p and NPY (n = 10); Comparisons between two groups were tested by t テスト
Knockdown of NPY abolishes the effects of inhibited miR-124-5p on hippocampal damages of depressed rats. a HE staining of hippocampal lesions of rats after inhibition of both miR-124-5p and NPY (n = 6); b ELISA of NE, 5-HT and DA expression in hippocampal tissues of rats after inhibition of both miR-124-5p and NPY (n = 6); C. Effect of inhibited NPY and suppressed miR-124-5p on SOD, MDA and GSH concentration in serum (n = 10); D. Effect of inhibited NPY and suppressed miR-124-5p on IL-1β, TNF-α and NO concentration in serum (n =10)。 Comparisons between two groups were tested by t テスト
Depression is a type of psychiatric disorder comprising a variety of conditions with diverse symptoms [20]. Though emerging studies have implied the role of miRNAs in depression, the precise action of miR-124-5p has been rarely investigated. Hence, the present study is projected for better comprehension of the mechanism of miR-124-5p/HDAC1/NPY axis in depression with the major outcome elaborating that silencing of either HDAC1 or miR-124-5p up-regulated NPY to improve memory and learning abilities of depressed rats.
To begin with, this study discovered HDAC1 expression in hippocampal tissues with the findings suggesting the elevation in the HDAC1 expression. Subsequently, with the purpose to decipher the functional roles of HDAC1 in depressed rats, loss-of-function assays were performed and it was disclosed that knockdown of HDAC1 increased the body weight, improved learning and memory abilities, attenuated pathological damage, up-regulated neurotransmitter expression, and suppressed oxidative stress and inflammation in depressed rats. Currently, a study has implied an increment in the expression of HDAC1 in depressive-like and anxiety-like phenotypes resulted by stress-offspring [21]. Moreover, the expression of HDAC1 is documented to up-regulate in penumbra in photothrombotic stroke [22]. Besides that, the incremental HDAC1 mRNA expression is found in granule and pyramidal cells in temporal lobe epilepsy [23]. As to the functional role of HDAC1, it has been reported that virus-mediated overexpression of neuronal HDAC1 in the hippocampus of mice imposes influences on loss of contextual fear memory in particular [24]. Mechanically, HDAC inhibitors reverse cognitive deficits found in neurodegenerative diseases and age-related memory loss [25]. Actually, it is accepted that the HDAC1 suppression by tianeptinaline has advanced neuroplasticity and reinforced memory [26]. As mentioned in a prior study, it is concluded that repression of HDAC1 inhibits the pathogenic processes that lead to motor neuron degeneration in mitochondrial diseases [27]. Experimentally, the silencing of HDAC1 by 5-thienyl-substituted 2-aminobenzamide-type is partially involved in the prevention of neuronal cell death in Parkinson's disease models [28]. Further supported by those researches, the protective effects of silenced HDAC1 have been witnessed in brain diseases, including but not limited to depression.
Then, our study discovered a targeting relationship between HDAC1 and miR-124-5p, which was supported by a prior study which suggests miR-124 transcription is in the charge of EVI1, acting by connection with the deacetylase HDAC1 [29]. miR-124-5p was the overexpressed gene in depressed rats and knockdown of miR-124-5p had the similar functions of silenced HDAC1 in depressed rats. In fact, there is a study indicating that the expression of miR-124 in the hippocampus is up-regulated from 5 to 6 weeks in depression-like behavior phenotypes [7]. Another study has identified the increase in the miR-124 expression in female with cocaine use disorder [30]. Also, it is previously described that miR-124-3p is highly expressed in stressed rodents in major depressive disorder [31]. Regarding to the effects, knockdown of miR-124 in the prefrontal cortex is reported to attenuate depression-like behavior of mice [9]. Besides that, miR-124 knockdown is believed to serve as an antidepressant agent of chronic corticosterone-induced gypenosides in mice [32]. Drawn from a prior study, knockdown of miR-124 can result in improved behavioral susceptibility to a milder stress paradigm [33]. It is reported that suppression of miR-124 by lentivirus transfection in the hippocampus can protect ketamine-induced neurodegeneration in vivo and in vitro [34]. Anyway, miR-124-5p suppression is an active actor to attenuate depression.
Furthermore, NPY expression was verified to be regulated by HDAC1 and miR-124-5p. The deteriorated deficits associated with HDAC2 in histone acetylation may be related to the decreased expression of NPY and can used to control anxiety-like and drinking behaviors [14]. NPY was down-regulated in depressed rats and up-regulation of NPY promoted learning and memory ability recovery in depressed rats. In the development of depressive-like behaviors, the rat models are manifested with reduced NPY expression [17]. Academically, the expression of NPY is evidenced to decline in mice with depression [35]. Consistently, the above-mentioned study findings are as same as the previous literature to some extent.
The novel findings of study suggested that inhibited miR-124-5p or suppressed HDAC1 attenuated learning and memory abilities and increased body weight of depressed rats. In addition, knockdown of miR-124-5p or inhibition of HDAC1 suppressed oxidative stress and inflammation and promoted neurotransmitter expression of depressed rats. Moreover, HDAC1 mediated miR-124-5p to regulate NPY. In the rescue experiments, knockdown of NPY abolished the protective effects of inhibited miR-124-5p on depressed rats.
In summary, this study highlighted the effect of HDAC1/miR-124-5p/NPY axis in depression with the major findings suggesting inhibited miR-124-5p or suppressed HDAC1 attenuated learning and memory abilities, increased body weight, suppressed oxidative stress and inflammation, as well as promoted neurotransmitter expression in depressed rats. HDAC1/miR-124-5p/NPY axis may provide a reference to treat neurological disorder, which may also update the existed knowledge of depression. However, further studies are still required for thorough comprehension of the complex mechanism of HDAC1/miR-124-5p/NPY axis in depression.
該当なし。
MicroRNA-124-5p
Histone deacetylase 1
Neuropeptide Y
Chronic unpredictable mild stress
Sprague–Dawley
Specific pathogen-free
Negative control
Open field test
Sucrose preference test
Sucrose preference percentage
Morris water maze test
Superoxide dismutase
Malondialchehyche
グルタチオン
Interleukin-β
Tumor necrosis factor-α
Nitric oxide
酵素免疫測定法
Hematoxylin–eosin
Bicinchoninic acid
Norepinephrine
Serotonin
ドーパミン
Reverse transcription quantitative polymerase chain reaction
Complementary DNA
ドデシル硫酸ナトリウム
Chromatin immunoprecipitation
Untranslated region
Mutant type
Wild type
一元配置分散分析はじめに
材料と方法
倫理声明
実験動物
ラットのグループ化とモデルの確立
行動機能テスト
標本コレクション
酸化ストレス関連および炎症性指数の検出
ヘマトキシリン-エオシン(HE)染色
神経伝達物質の発現検出
逆転写定量的ポリメラーゼ連鎖反応(RT-qPCR)
ウエスタンブロット分析
クロマチン免疫沈降(ChIP)アッセイ
デュアルルシフェラーゼレポーター遺伝子アッセイ
統計分析
結果
HDAC1またはmiR-124-5pのいずれかを阻害すると、うつ病のラットの体重が増加します
HDAC1またはmiR-124-5pのいずれかを阻害すると、うつ病のラットの学習能力と記憶能力が向上します
HDAC1またはmiR-124-5pのいずれかを阻害すると、うつ病のラットの病理学的ニューロン損傷が軽減されます
HDAC1またはmiR-124-5pのいずれかの阻害-うつ病ラットにおける神経伝達物質の発現をアップレギュレートします
HDAC1またはmiR-124-5pのいずれかを阻害すると、うつ病のラットの酸化ストレスと炎症が抑制されます
HDAC1はmiR-124-5pを仲介してNPYを規制します
Knockdown of NPY Abolishes the Protective Effects of Inhibited miR-124-5p on Depressed Rats


Discussion
Conclusion
データと資料の可用性
略語
ナノマテリアル
- tinyMLと自動機械学習を使用した効果的なIoTアプリケーションの構築
- ST:低コストのSO-8パッケージに豊富なアナログとDMAを備えた8ビットMCU
- 粘り強さと忍耐力でリードする
- 人工ニューロンによる教師なし学習
- 機械学習で信頼性を高め、メンテナンスの成果を向上させる
- 高度な農薬活性を備えたスマートナノマテリアルおよびナノコンポジット
- 自己整流スイッチ特性を備えたTaO / HfOxおよびTaO / AlO xRRAMの保持モデル
- CuSナノ粒子でコーティングされた着色および導電性CuSCN複合材料の容易な合成
- RGOと3次元グラフェンネットワークが高性能でTIMを共同修正
- 窒化処理を施したHfO2ベースのRRAMの伝導メカニズムと耐久性の向上
- 抵抗変化型メモリのモデリングとシミュレーションに関する集合的研究