


ヒト臍帯間葉系幹細胞由来のエクソソームマイクロRNA-18b-3pは、LEPを標的とすることにより子癇前症の発生を阻害します
要約
マイクロRNAを発現するヒト臍帯間葉系幹細胞(hucMSC)に由来するエクソソームは、ヒトの疾患で強調されています。ただし、子癇前症(PE)のhucMSCs由来のエクソソームmiR-18b-3pの詳細な分子メカニズムはさらなる調査が残っています。 PEの発生に対するエクソソームとmiR-18b-3p /レプチン(LEP)の影響を調査することを目的とした。 hucMSCおよびhucMSC-エクソソーム(Exos)の形態が特定されました。エクソソームをmiR-18b-3pを発現するさまざまなレンチウイルスに感染させて、PEにおけるmiR-18b-3pの役割を調べました。 PEラットモデルは、 N の腹腔内注射によって確立されました。 -ニトロ-1-アルギニンメチルエステル。 LEPおよびmiR-18b-3pの発現は、PEラット胎盤組織でテストされました。また、LEPおよびmiR-18b-3pの発現に対するエクソソームの影響が検出されました。収縮期血圧(SBP)、タンパク尿、炎症性因子、胎児ラットと胎盤の重量、およびPEラットの細胞アポトーシスが検出されました。最後に、miR-18b-3pとLEPの関係は、デュアルルシフェラーゼレポーター遺伝子アッセイとRNAプルダウンアッセイを使用して検証されました。エクソソーム、miR-18b-3pの回復、またはLEPの阻害により、PEラットのSBPおよびタンパク尿が減少し、胎児ラットおよび胎盤の重量が増加し、炎症性因子の血清レベルが減少し、PEラットのアポトーシス細胞が抑制され、抑制効果が発揮されました。 PEの進行について。 PEラットの胎盤組織ではmiR-18b-3pが減少し、LEPが増加しました。 LEPはmiR-18b-3pの直接の標的遺伝子でした。 miR-18b-3pのアップレギュレーションまたはエクソソームの治療は、LEPの発現を抑制し、PEの発生を減少させましたが、miR-18b-3pのダウンレギュレーションは逆の効果をもたらしました。ダウンレギュレーションされたLEPは、PEラットに対するmiR-18b-3pの減少の影響を逆転させました。 HucMSCs由来のエクソソームmiR-18b-3pは、LEPを標的にしてPEの発生と発達に関与します。この研究は、PEのメカニズムと調査のための新しい理論的基礎を提供するかもしれません。
はじめに
タンパク尿と高血圧を特徴とする子癇前症(PE)[1]は、ヒトの妊娠における胎児と妊産婦の死亡率と罹患率の主な原因です[2]。 PEの病因と病因は明らかではなく[3]、これは異常な栄養膜浸潤に関連し、母体の内皮機能障害、慢性胎盤灌流、高血圧を引き起こし、有害な結果をもたらすことが報告されています[4]。胎児および胎盤への送達を除いて、PEに対する特定の治療法はありません[5]。したがって、この疾患の予後を改善するための治療標的を探索することが急務です。
ヒト臍帯(huc)は、さまざまな栄養因子やサイトカインを分泌するだけでなく、強力な抗炎症作用や免疫調節作用を示す間葉系幹細胞(MSC)の適切な供給源です[6]。ある研究では、PEが臍帯ウォートンゼリー由来のMSCにおける神経膠マーカーの発現を促進することが確認されています[7]。 PEラットの胎盤形態および血管新生に対するhucMSCエクソソーム(Exos)の保護効果も報告されています[8]。エクソソームは小さな(50〜100 nm)分泌小胞であり、循環によって発がん性因子をカプセル化して離れた場所や周囲の細胞に伝達することにより、腫瘍微小環境内の細胞間のコミュニケーションを仲介します[9]。ある研究では、エクソソームによるPE患者の内皮細胞へのsFlt-1とsEngの効果的な移行によって誘発される血管機能と合併症の損傷が明らかになりました[10]。マイクロRNA(miRNA)は、18〜25ヌクレオチドの長さの内因性の非コードRNAであり、転写後のレベルで遺伝子の発現を調節します[11]。ある研究のデータは、miR-18bの発現が、PEにおける細胞浸潤、生存率、および栄養膜細胞の移動に影響を与えることを報告しています[12]。さらに、ウー等。 miR-18bは、高グルコースによって誘発されるヒト網膜内皮細胞の増殖を弱めることを提案しました。これは、糖尿病性網膜症の病因のメカニズムを理解するための新しい洞察を提供する可能性があります[13]。ただし、PEにおけるhucMSC由来のエクソソームmiR-18b-3pの役割は不明なままです。レプチン(LEP)は、細胞の分化/増殖および生理学的状態の免疫に多面的な効果をもたらし、胎盤を含む他の組織に加えて、主に脂肪細胞から出現します[14]。ある研究では、異常なLEPプロモーターのメチル化がPEの進行に関与していることが確認されています[15]。別の研究では、胎盤が妊娠中のLEPの発現の主要部位であることが示唆されています[16]。それにもかかわらず、miR-18b-3pとLEPの間の結合関係はまだとらえどころのないです。したがって、LEPが関与するPEにおけるhucMSC由来のエキソソームmiR-18b-3pの役割を調査することを目的とし、hucMSC由来のエキソソームmiR-18b-3pがLEPを標的とすることでPEの進行を阻害する可能性があると推測しました。
>材料と方法
倫理的承認
この研究は、武漢大学人民病院の倫理委員会によって承認されました。すべての参加者はインフォームドコンセントの文書に署名しました。すべての動物実験は、武漢大学人民病院の国際委員会による実験動物の管理と使用に関するガイドと集計されました。
HucMSCの分離、文化、および識別
健康な子宮頸管によって送達された胎児の臍帯を収集し、ミンチに切断し、ふるいで濾過し、次にリン酸緩衝生理食塩水(PBS)溶液と混合した。臍帯組織を1500r / minで5分間、遠心半径10cmで遠心分離しました。組織を10%ウシ胎児血清(FBS)を含むダルベッコ改変イーグル培地(DMEM)/ F12で懸濁し、培養フラスコに移しました。液体は4日後に交換され、その後3日ごとに交換されました。コンフルエンスが約90%に達したときに細胞を継代培養した。 hucMSCの付着成長と形態を光学顕微鏡で観察した。細胞をオイルレッドO染色液(Beyotime Institute of Biotechnology、上海、中国)で染色してhucMSCの骨形成分化を検出し、アルカリホスファターゼ(ALP)染色液(Beyotime)で染色してhucMSCの脂肪生成分化を検出しました。フローサイトメーター(Beckman Coulter Life Sciences、Brea、CA、USA)を採用して、CD73、CD166(1:10、BD Biosciences、Franklin Lakes、NJ、USA)およびCD105(1:20、AbD Serotec、Oxford、英国)。
HucMSC-Exosの抽出と識別
よく成長しているhucMSCを培養した。上澄みを収集し、28,500 r / minで1時間、遠心分離半径10cmで遠心分離しました。上清を捨て、細胞を2%グルタルアルデヒドと1%オスミウム酸で固定し、エタノールで脱水し、酸化プロピレンに浸し、2時間乾燥させ、Epon812で包埋し、スライスしました。スライスはそれぞれウランと鉛で染色されました。最後に、エクソソームを電子顕微鏡で観察しました。 Nanosight検出器(Malvern Instruments、Malvern、UK)を使用して、エクソソームナノ粒子とそのサイズのブラウン運動イメージングを検出しました。 hucMSC-Exosの表面マーカーはウエスタンブロットアッセイによって同定され、その結果は、hucMSC-ExosがCD9、CD81、およびCD63を発現することを示しました。
レンチウイルス感染法
HucMSCは、miR-18b-3pベクターの低発現とmiR-18b-3pベクターネガティブコントロール(NC)の低発現を含むレンチウイルスに感染しました(Shanghai GenePharma Co、Ltd、Shanghai、China)。最後に、安定して発現されたhucMSC-antagomirNCおよびhucMSC-miR-18b-3pantagomirが得られた。細胞を48時間培養し、上清を回収して超遠心分離で遠心分離し、対応するExos-antagomirNCおよびExos-miR-18b-3pantagomirを取得しました。
実験動物
健康状態が良好で性的に成熟しているウィスターラット(体重200〜250 g、8 wの年齢、性別を問わない)を選択しました(武漢大学実験動物センター、武漢、中国)。ラットは、18〜28°Cの温度、40〜70%の相対湿度、適切な食餌と水を備えたバリアシステムで飼育されました。
ラットPEモデルの確立
ラットPEモデルは、記事[17]を参照して、50 mg / kgの一酸化窒素シンテターゼ阻害剤であるN(G)-ニトロ-1-アルギニンメチルエステル(L-NAME、Beyotime)の腹腔内注射によって確立されました。 PEモデルの確立に成功したのは、20mmgHgおよび115mmHgを超える血圧の上昇と、タンパク尿の増強に基づいていました。
動物のグループ化
メスのラットとオスのラットは1:1でランダムに同棲し、2匹のラットは午後5時から6時まで個別の特別なケージに入れられました。その前日。翌日、雌ラットの膣分泌物中の精子を膣栓と顕微鏡で観察した。結果が同時に陽性であった場合、その日は妊娠0日として記録されました。 13日から 妊娠日、ラットを6つのグループ(各グループに10匹のラット)に分けました:正常グループ(妊娠13日から20日まで同量の正常生理食塩水を腹腔内注射)、PEグループ(L-NAME [50 mg / [kg /日]は妊娠13日から20日まで腹腔内注射され、20μLの通常の生理食塩水が妊娠16日から19日まで胎盤に注射されました)、PE + miR-NCグループ(L-NAME [50 mg / kg /日]妊娠13日目から20日目まで腹腔内注射し、妊娠16日目から19日目までに20μLの4 nmol miR-NCを胎盤に注射しました)、PE + miR-18b-3pアゴミールグループ(L-NAME [50 mg / kg /日]は妊娠13日から20日まで腹腔内注射され、20μLの4 nmol miR-18b-3pアゴミールが妊娠16日から19日目に胎盤に注射されました) 、PE + miR-18b-3pアンタゴミールグループ(L-NAME [50 mg / kg /日]を妊娠13日目から20日目まで腹腔内注射し、20μLの4 nmolmiR-18b-3pアンタゴミールをp妊娠16日目から19日目までの胎盤)、PE + miR-18b-3pアンタゴミール+小干渉RNA(si)-LEPグループ(L-NAME [50 mg / kg /日]を13日目から20日目まで腹腔内注射妊娠期間中、および20μLの4 nmol miR-18b-3pアンタゴミールとsi-LEPを妊娠16日目から19日目に胎盤に注射しました)およびPE + si-LEPグループ(L-NAME [50 mg / kg per ]は妊娠13日から20日まで腹腔内注射され、20μLの4 nmol si-LEPが妊娠16日から19日まで胎盤に注射されました)。ラットは、レンチウイルスを運ぶエクソソームとエクソソームで治療されました。ラットは5つのグループ(各グループに10匹のラット)に割り当てられました:正常グループ(妊娠13日から20日まで同量の正常生理食塩水が腹腔内注射された)、PEグループ(L-NAME(50 mg / kg /日) )妊娠13日目から20日目まで腹腔内注射し、妊娠16日目から19日目まで20μLの通常生理食塩水を胎盤に注射しました)、PE + Exosグループ(L-NAME(50 mg / kg /日)を妊娠13日から20日まで腹腔内注射し、20μLのExos(80μgのエキソソームを20μLの通常の生理食塩水に懸濁)を妊娠16日から19日目に胎盤に注射しました)、PE + Exos-antagomir NCグループ(L-NAME(50 mg / kg /日)を妊娠13日から20日まで腹腔内注射し、20μLのExos-antagomir NC(80μgのエキソソームを20μLの通常の生理食塩水に懸濁)を胎盤に注射しました。妊娠16日目から19日目)およびPE + Exos-miR-18b-3pアンタゴミールグループ(L-NAME(50 mg / kg /日)を13日目から腹腔内注射)妊娠20日目までに20μLのExos-miR-18b-3pアンタゴミール(80μgのエクソソームを20μLの生理食塩水に懸濁)を妊娠16日目から19日目に胎盤に注射しました。
収縮期血圧(SBP)の検出と24時間タンパク尿の測定
ラットの圧力は、ラットの尾動脈血圧測定によって測定された。妊娠10、13、16、19日目に、ラット尾動脈圧検出器(Tensys(R)Medical Inc.、San Diego、CA、USA)を使用して、すべての妊娠ラットの尾カフSBPを測定しました。圧力は短時間で3回測定されました。次に、平均値を血圧としました。
自由食と水を与えた場合、妊娠10、13、16、19日目に妊娠ラットの24時間尿を採取し、武漢大学人民病院腎臓内科でタンパク質含有量を検出した。
サンプルコレクション
妊娠21日目に妊娠ラットを3%ペントバルビタールナトリウムで麻酔した。ラットの末梢血を保存し、遠心分離して血清を採取し、–20°Cの冷蔵庫に保管して待機しました。次に、帝王切開でラット胎児と胎盤を採取し、卵膜と接続された臍帯を取り除き、ラット胎児に接続された臍帯を切断した。胎盤および胎児ラットを無菌ガーゼに置いて、それぞれ血液および羊水を乾燥させ、次に分析天秤に載せて重量を秤量した。胎盤組織の一部を4%パラホルムアルデヒドで固定し、エタノールで脱水し、キシレンで透明にし、パラフィンで包埋し、ヘマトキシリン-エオシン(HE)染色および末端デオキシヌクレオチドトランスフェラーゼ媒介デオキシウリジン三リン酸-ビオチンのために連続的にクロススライス(5μm)しましたニックエンドラベリング(TUNEL)染色。残りは、逆転写定量的ポリメラーゼ連鎖反応(RT-qPCR)検出、ウエスタンブロット分析、および酵素結合免疫吸着測定法(ELISA)のために-80°Cで保存されました。
ELISA
血清中の腫瘍壊死因子-α(TNF-α)、インターロイキン(IL)-1βおよびIL-6の含有量をELISAで試験しました。 TNF-α、IL-1βおよびIL-6の濃度は、キット(R&D Systems、ミネソタ州ミネアポリス、米国)の指示に従って決定された。光学密度(OD)値(490 nm)は、マイクロプレートリーダー(Thermo Fisher Scientific、マサチューセッツ州、米国)によってテストされました。対応する検量線は、横軸にOD値、縦軸に対応する標準サンプルの濃度を使用して得られました。 TNF-α、IL-1β、IL-6の濃度は検量線から計算されました。
HE染色
胎盤組織のパラフィンサンプルをキシレンで清澄化し、従来の勾配アルコールで脱水し、ヘマトキシリンで染色し、1%塩酸アルコールで区別し、1%アンモニア水で青色に戻しました。次に、組織を1%エオシン溶液で対比染色し、脱水し(それぞれ、75%、90%、95%エタノール、無水エチルアルコール)、キシレンで透明にし、乾燥させ、ブロックし、電子顕微鏡で観察しました。
TUNEL染色
パラフィン包埋切片は、指示に従って定期的に脱ロウおよび脱水し、その後、TUNELキット(Nanjing Kejin Biotechnology Co.、Ltd.、Jiangsu、China)によってアポトーシスを検出しました。 4,6-ジアミノ-2-フェニルインドール(Shanghai Baitai Biotechnology Co.、Ltd.、Shanghai、China)を使用して、蛍光顕微鏡(Nikon、Tokyo、Japan)を使用してTUNEL陽性細胞を観察しました[18]。
RT-qPCR
胎盤組織の重さを量った。 50〜100 mgあたりの胎盤組織に、1 mLのTRIzol(Invitrogen、米国カリフォルニア州カールズバッド)を追加し、完全に溶解しました。組織に200μLのクロロホルムを添加し、4°C、12,000rpmで遠心分離してトータルRNAを抽出しました。 RNAの濃度と純度はDU-800タンパク質核酸分光光度計(Beckman)によって決定されました。 U6およびβ-アクチンをローディングコントロールとして利用しました。 PCRプライマーは、Shanghai Sangon Biotechnology Co. Ltd.(Shanghai、China)によって設計および配合されました。プライマー配列を表1に示します。RNA逆転写キット(Sangon)の説明書に基づいて、RNAをcDNAに逆転写しました。 PCRを増幅し、生成物をアガロースゲル電気泳動で確認しました。データは2 -ΔΔCt によって計算されました メソッド。
<図>ウエスタンブロットアッセイ
胎盤組織の総タンパク質は、放射性免疫沈降アッセイ細胞溶解バッファー(Beyotime)によって抽出されました。 HucMSC-Exoを使用してバッファーを抽出し、14,000rpmで遠心分離しました。血清中のエクソソームマーカータンパク質(CD81、CD63およびCD9)のタンパク質発現をテストするために、上清を保存しました。タンパク質濃度は、ビシンコニン酸キット(Beyotime、P0010)によって決定されました。タンパク質の定量結果に従ってサンプルをロードし、10%ドデシル硫酸ナトリウム-ポリアクリルアミドゲル電気泳動で処理し、メンブレンに転写しました。メンブレンを5%スキムミルクでブロックし、一次抗体LEP、CD63、CD81、CD9、およびβ-アクチン(4 mL、1:1000、Santa Cruz Biotechnology、Inc、Santa Cruz、CA、USA)でプローブし、再プローブしました。 4mLの二次抗体ヤギ抗ウサギIgG /西洋ワサビペルオキシダーゼを用いて、曝露および開発しました。 β-アクチンを内部参照として利用しました。グレー値は、ゲルグラフィック分析ソフトウェアImageLabによって分析されました。
デュアルルシフェラーゼレポーター遺伝子アッセイ
オンライン予測ソフトウェアhttps://cm.jefferson.edu/を採用して、miR-18b-3pとLEPのターゲット関係、およびmiR-18b-3pとLEP 3 '非翻訳領域(UTR)の結合部位を予測しました。 )。 miR-18b-3p結合部位を含むLEP3'UTRプロモーター領域配列を構成した。 LEP 3'UTR野生型(WT)プラスミドとLEP 3'UTR変異型(MUT)を構築しました。組換えプラスミドは、それぞれLEP3'UTR-WTおよびLEP3'UTR-MUTと名付けられました。培養された293T細胞は、miR-18b-3pミミックおよびLEP 3'UTR-WT、miR-18b-3pミミックおよびLEP 3'UTR-MUT、ミミックNCおよびLEP 3'UTR-WT、ミミックNCおよびLEP3'UTR-30時間MUT。次に、293T細胞を収集しました。細胞内のホタルおよびレニラルシフェラーゼ活性は、デュアルルシフェラーゼレポーター遺伝子検出キット(Promega、ウィスコンシン州マディソン、米国)に準拠した発光測定によって測定されました。
RNAプルダウンアッセイ
ビオチン化RNAプローブ(Bio-miR-NC、Bio-miR-18b-3pおよびBio-miR-18b-3p-Mut)を293T細胞の溶解物とインキュベートし、抗生物質ストレプトマイシンと結合した磁気ビーズを使用して抽出しました。実験は、Pierce磁気RNAプルダウンキット(Pierce、IL、USA)の指示に基づいて実施されました。 RNAは、TRIzol(Pierce)を使用して溶出および精製しました。 RNA複合体におけるLEPの濃縮は、前述のようにRT-qPCRを使用して定量化されました[19]。
統計分析
すべてのデータは、SPSS 21.0ソフトウェア(IBM Corp. Armonk、NY、USA)によって説明されました。測定データは、平均±標準偏差として示されました。データは独立したサンプル t によって実施されました 2つのグループの比較をテストし、複数のグループ間の比較は、一元配置分散分析(ANOVA)とそれに続くテューキーの事後検定によって評価されました。統計的有意性の基準は p に設定されました <0.05。
結果
HucMSCおよびHucMSC-Exosの形態と識別
臍帯組織塊を倒立顕微鏡下で観察した。 3日目に細胞が組織塊から這い出たことがわかりました。細胞は紡錘形と糸状を示し、約5日でコロニーのように成長しました。 3継代まで培養した場合、細胞の形態は均一で長い紡錘状であり、線維芽細胞の形態に類似しており、配列は規則的でした(図1a)。 hucMSCの脂肪生成分化の2週間後、脂肪滴が細胞質に形成され、脂肪滴は倒立顕微鏡下でクランツ構造を示し(図1b)、単離された培養hucMSCが脂肪生成分化の能力を持っていることを示唆しています。 2週間の骨形成分化後、倒立顕微鏡下で多数の茶色のカルシウム結節が見られ(図1c)、単離された培養hucMSCが骨形成分化の能力を持っていたことを示しています。細胞の免疫表現型をテストするためにフローサイトメーターが採用され、その結果には、細胞がMSCの表面マーカーCD73、CD105、およびCD166を過剰発現したことが含まれていました(図1d)。
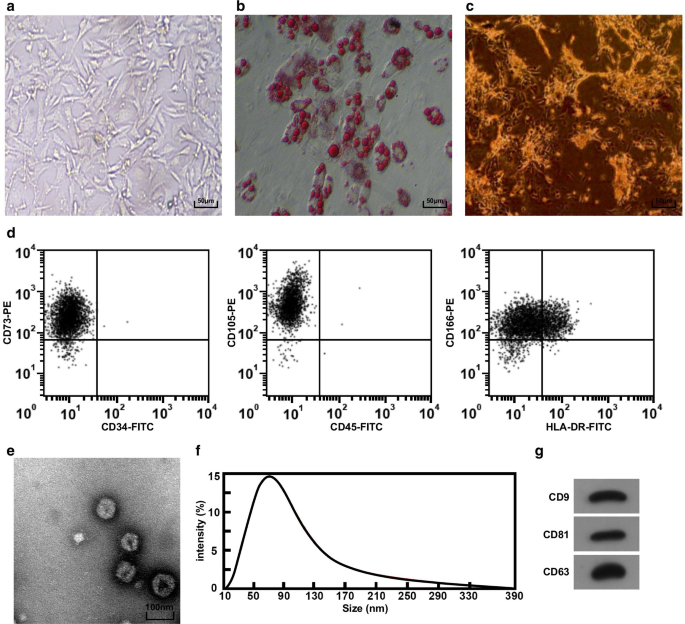
hucMSCおよびhucMSC-Exosの形態と同定。 a 倒立顕微鏡下でのhucMSCの形態 b hucMSCはオイルレッドO染色によってテストされました。 c hucMSCはALP染色によってテストされました。 d フローサイトメトリーを使用して免疫表現型を検出した。 e TEMで観察されたhucMSC-Exosの形状とサイズ。 f Nanosight分析を使用したエクソソームの粒子サイズ分布の検出。 g hucMSC-ExosにおけるCD9、CD81、およびCD63のタンパク質発現は、ウエスタンブロットアッセイによって検出されました
hucMSC-Exosの形態は、TEMによって観察され、その結果、エクソソームは円形または楕円形であり、中心密度が低く、両側に濃い染色が見られました(図1e)。ナノサイト分析を利用してエクソソームの粒子サイズを分析した結果、粒子サイズは主に40〜100 nmに分布し、80 nm付近に集中していることがわかりました(図1f)。ウエスタンブロットアッセイにより、すべての表面マーカーCD81、CD63、およびCD9がhucMSC-Exosで発現していることが明らかになりました(図1g)。
miR-18b-3pを復元すると、PEラットの病理学的特徴が緩和されます
SBPと24時間タンパク尿の結果は次のことを示しました:投与前(妊娠10日目)の6群でSBPと24時間タンパク尿に有意差はありませんでした。妊娠19日目のSBPおよび24時間タンパク尿は、正常ラットで明らかな違いを示さなかった。 miR-NC、miR-18b-3pアゴミール、miR-18b-3pアンタゴミール、miR-18b-3pアンタゴミール+ si-LEP、si-NCまたはsi-LEP、SBPおよび24時間で治療されたPEラットまたはPEラットタンパク尿は妊娠13日目に増加し始めました。 miR-18b-3pアゴミールとsi-LEPで治療したPEラットでは、妊娠16日目と19日目でSBPと24時間タンパク尿に明確な違いはありませんでした。 PEラットは、妊娠19日目にSBPと24時間タンパク尿が増加した。この増加は、miR-18b-3pの上昇によって減少しましたが、miR-18b-3pの阻害によってさらに強化されました。 LEPの低下により、PEラットの妊娠19日目のSBPおよび24時間タンパク尿におけるmiR-18b-3pのダウンレギュレーションの役割が無効になりました(図2a、b)。
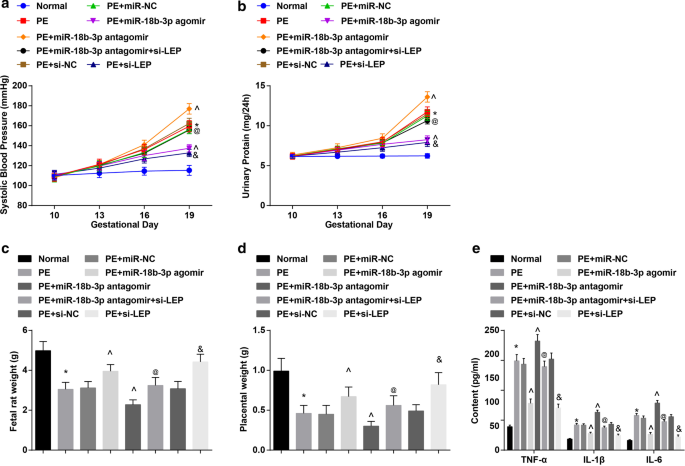
miR-18b-3pを復元すると、PEラットの病理学的特徴が緩和されます。 a ラットにおけるSBPの結果。 b ラットにおける24時間タンパク尿の結果。 c 胎児ラットの体重変化。 d ラットの胎盤重量の変化。 e 血清中の炎症性因子の変化は、ELISAを使用して検出されました。 n =10、* p <0.05対正常群。 ^ p <0.05対PE + miR-NCグループ。 @ p <0.05対PE + miR-18b-3pアンタゴミールグループ。 & p <0.05対PE + si-NCグループ。測定データは平均±標準偏差として表され、複数のグループ間の比較は一元配置分散分析とそれに続くテューキーの検定によって評価されました
PEラットでは胎児ラットと胎盤の体重が減少した。ダウンレギュレーションされたmiR-18b-3pまたはダウンレギュレーションされたLEPは増加しましたが、ダウンレギュレーションされたmiR-18b-3pは、PEラットの胎児ラットと胎盤の体重を減少させました。 LEPサイレンシングは、PEラットの胎児ラットと胎盤の体重に対するmiR-18b-3p阻害の効果を逆転させました(図2c、d)。
PEラットの血清中の炎症性因子が検出された。 PEラットではTNF-α、IL-1β、IL-6の含有量が増加することがわかりました。 miR-18b-3pの上昇またはLEP阻害は抑制され、miR-18b-3pの低下はTNF-α、IL-1βおよびIL-6の含有量を促進しました。阻害されたmiR-18b-3pがTNF-α、IL-1β、およびIL-6の含有量に及ぼす影響は、LEPの枯渇によって無効になりました(図2e)。
過剰発現したmiR-18b-3pは、PEラットの胎盤組織の組織病理学的変化を改善します
正常なラットでは、胎盤絨毛は血管が豊富で明確な構造を持っており、合胞体栄養膜が胎盤絨毛の主要な栄養膜であり、細胞栄養芽層は少なかった。 miR-NC、miR-18b-3p antagomir、si-NCまたはmiR-18b-3p antagomir + si-LEPで治療されたPEラットまたはPEラットでは、胎盤絨毛の数が減少し、構造がぼやけて萎縮し、一部の絨毛がフィブリノイド壊死が行われ、胎盤絨毛の合胞体栄養膜結節の数が増加し、ほとんどの絨毛は未成熟でした。 miR-18b-3pアゴミールとsi-LEPで治療したPEラットでは、栄養細胞の数が減少し、病理学的変化が緩和されました(図3a)。
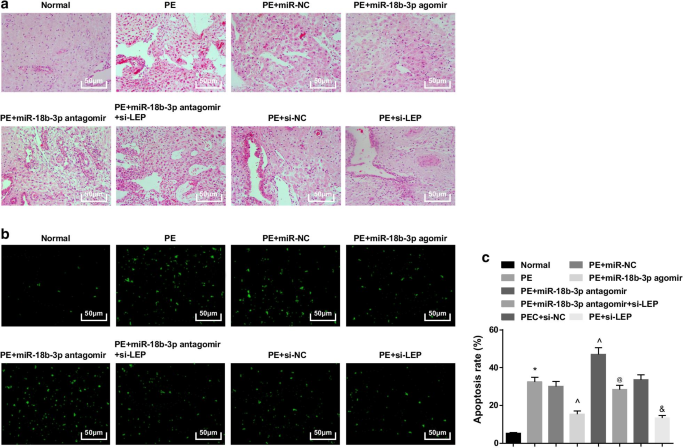
過剰発現したmiR-18b-3pは、PEラットの病理学的変化を改善し、胎盤組織のアポトーシス細胞を抑制します。 a HE染色を利用して、胎盤組織の病理学的特徴を試験した。 b TUNEL染色は、PEラットの胎盤組織のアポトーシス細胞を決定するために実施されました。 c 細胞アポトーシス率はTUNEL染色によって検出されました。 n =10、* p <0.05対正常群。 ^ p <0.05対miR-NCグループ。 @ p <0.05対miR-18b-3pアンタゴミールグループ。 & p <0.05対si-NCグループ。測定データは平均±標準偏差として表され、複数のグループ間の比較は一元配置分散分析とそれに続くテューキーの検定によって評価されました
TUNEL染色は、少数のアポトーシス細胞が見られる可能性があることを示唆しました。 PEラットはアポトーシス細胞が増加し、miR-18b-3pの上昇とLEPサイレンシングによって減少し、miR-18b-3pの阻害によってさらに増強されました。 LEPサイレンシングは、PEラットのアポトーシス細胞数に対するmiR-18b-3p阻害の影響も逆転させました(図3b、c)。
まとめると、miR-18b-3pが上方制御されている、またはLEPが抑制されているラットは、組織学的にPEの進行度が低下しており、LEPが沈黙していると、抑制されたmiR-18b-3pの治療効果が失われる可能性があります。
miR-18b-3pはダウンレギュレーションされていますが、PEラット胎盤組織およびmiR-18b-3pターゲットLEPではLEPがアップレギュレーションされています
上記の結果に基づいて、LEPのダウンレギュレーションは、病理学および組織学におけるPEラットに対するmiR-18b-3pのダウンレギュレーションの治療効果を逆転させました。したがって、miR-18b-3pがLEPに関連している可能性があると仮定しました。
ウエスタンブロットアッセイとRT-qPCRにより、PEラットはmiR-18b-3pが減少し、LEP発現レベルが増加したことが明らかになりました。 miR-18b-3pアゴミールの治療はPEラットでmiR-18b-3pをアップレギュレーションし、LEPをダウンレギュレーションしましたが、miR-18b-3pアンタゴミールの治療はLEP発現を増加させました。 LEPサイレンシングは、PEラットのLEP発現に対するmiR-18b-3p減少の促進効果を逆転させました(図4a–c)。
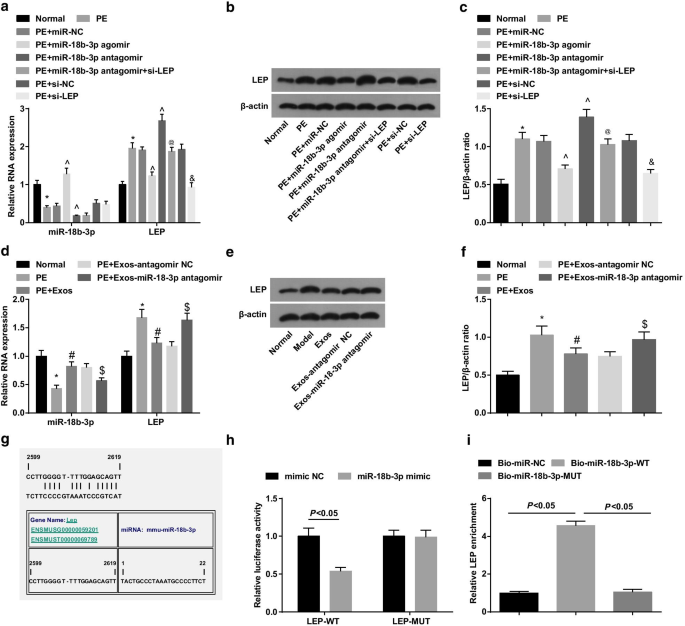
PEラットの胎盤組織ではmiR-18b-3pがダウンレギュレーションされ、LEPがアップレギュレーションされます。 a 胎盤組織におけるmiR-18b-3pおよびLEPmRNAの発現は、RT-qPCRを使用して検出されました。 b 胎盤組織におけるLEPのタンパク質バンド。 c 胎盤組織におけるLEPタンパク質発現を検出するためにウエスタンブロットアッセイを実施した。 d エクソソーム処理後の胎盤組織におけるmiR-18b-3pおよびLEPmRNAの発現は、RT-qPCRを使用して検出されました。 e エクソソーム処理後の胎盤組織におけるLEPのタンパク質バンド。 f エクソソーム処理後のLEPタンパク質発現を検出するためにウエスタンブロットアッセイを実施した。 g オンラインソフトウェアによって予測されるmiR-18b-3pとLEPの結合部位。 h miR-18b-3pとLEPのターゲット関係は、デュアルルシフェラーゼレポーター遺伝子アッセイによって検証されました。 i RNAプルダウンアッセイによって検証されたmiR-18b-3pとLEPの間のターゲティング関係。 n =10、* p <0.05対正常群。 ^ p <0.05対miR-NCグループ。 @ p <0.05対miR-18b-3pアンタゴミールグループ。 & p <0.05対si-NCグループ。 # p <0.05対PEグループ。 $ p <0.05対PE + Exos-antagomirNCグループ。測定データは平均±標準偏差として表され、複数のグループ間の比較は一元配置分散分析とそれに続くテューキーの検定によって評価されました
ウエスタンブロットアッセイとRT-qPCRを使用して、PEラットにおけるエクソソームの役割を調べました。結果は、PEラットにおいてエクソソームがmiR-18b-3pをアップレギュレーションし、LEPをダウンレギュレーションしたことを示しており、PE発生に対するエクソソームの抑制効果を示しています。さらに、miR-18b-3pアンタゴミールを運ぶエクソソームは、PEラットでmiR-18b-3pのダウンレギュレーションとLEPのアップレギュレーションを誘発しました(図4d–f)。
miR-18b-3pとLEPのターゲット関係は、バイオインフォマティクスのオンライン予測ソフトウェアhttps://cm.jefferson.edu/によって予測されました(図4g)。デュアルルシフェラーゼレポーター遺伝子アッセイは、miR-18b-3pミミックがLEP 3'UTR-WTのルシフェラーゼ活性を低下させたが、LEP 3'UTR-MUTのルシフェラーゼ活性には影響を与えなかったことを示唆しました(図4h)。さらに、RNAプルダウンアッセイにより、WT-ビオチン化miR-18b-3pによってLEP濃縮が増加することが明らかになりました(図4i)。これらの発見は、LEPがmiR-18b-3pの標的遺伝子であることを示しています。
hucMSC-ExosはPEラットの病理学的特徴を弱める
SBPと24時間の結果は、投与前(妊娠10日目)の5つのグループでSBPと24時間のタンパク尿に有意差がないことを示しました。 SBP and 24-h proteinuria in day 19 of gestation showed no distinct difference in normal rats. In PE rats, SBP and 24-h proteinuria began to raise at day 13 of gestation. There was no distinct difference of SBP and 24-h proteinuria in day 16 and day 19 of gestation in PE rats treated with hucMSC-Exos and hucMSC-Exos transmitting antagomir NC. SBP and 24-h proteinuria heightened in day 19 of gestation in the PE rats, while the increase was reduced by injection of hucMSC-Exos. Inhibiting miR-18b-3p reversed the effect of hucMSC-Exos on SBP and 24-h proteinuria in day 19 of gestation in PE rats (Fig. 5a, b).

hucMSC-Exos attenuate pathological characteristics of PE rats. a Results of SBP in rats after exosome treatment. b Results of 24-h proteinuria in rats after exosome treatment. c Weight changes of fetal rats after exosome treatment. d Changes of placental weight in rats after exosome treatment. e Changes of inflammation factors after exosome treatment in serum were determined using ELISA. n = 10, *p < 0.05 versus the normal group. # p < 0.05 versus the PE group. $ p < 0.05 versus the PE + Exos-antagomir NC group. Measurement data were depicted as mean ± standard deviation, and comparisons among multiple groups were assessed by one-way ANOVA followed by Tukey’s test
The weight of fetal rat and placenta was measured, and we found that the PE rats had decreased weight of fetal rat and placenta; miR-18b-3p downregulation abolished the role of hucMSC-Exos in the weight of fetal rat and placenta in PE rats (Fig. 5c, d).
Inflammatory factors in serum were detected using ELISA. TNF-α, IL-1β and IL-6 contents remarkably increased in PE rats. Exosomes treatment decreased TNF-α, IL-1β and IL-6 contents in serum of PE rats, which were enhanced by injection of exosomes inhibiting miR-18b-3p (Fig. 5e).
Exosomes Alleviates Pathological Change and Inhibit Apoptosis of Placenta Tissues of PE Rats
In normal rats, the placental villus was abundant in blood vessels with a clear structure, syncytiotrophoblasts were the main trophoblast in placental villi, and there were fewer cytotrophoblasts. In the PE rats and PE rats treated with hucMSC-Exos-miR-18b-3p-antagomir, the number of placental villi reduced, the structure was blurred and atrophied, some villi were presented fibrinoid necrosis, and the number of syncytiotrophoblast nodules in placental villi enhanced, and most of the villi were immature. The pathological change was improved in the PE rats treated with hucMSC-Exos or hucMSC-Exos-antagomir NC versus the PE rats and PE rats treated with hucMSC-Exos-miR-18b-3p antagomir (Fig. 6a).

Exosomes alleviate pathological change and decrease apoptotic cells of placenta tissues in PE rats. a HE staining was utilized to test pathological features of placenta tissues in PE rats after exosome treatment. b TUNEL staining was implemented to determine apoptotic cells of placenta tissues in PE rats after exosome treatment. c Cell apoptosis rate was detected by TUNEL staining. n = 10, *p < 0.05 versus the normal group. # p < 0.05 versus the PE group. $ p < 0.05 versus the PE + Exos-antagomir NC group. Measurement data were depicted as mean ± standard deviation, and comparisons among multiple groups were assessed by one-way ANOVA followed by Tukey’s test
TUNEL staining indicated that in normal rats, a small number of apoptotic cells could be seen. PE rats had enhanced apoptotic cells, and reduced miR-18b-3p reversed the impacts of hucMSC-Exos on the number of apoptotic cells in placenta tissues from PE rats (Fig. 6b, c).
ディスカッション
PE is a multisystem pregnancy disorder characterized by proteinuria and either high blood pressure or other adverse conditions and is linked to a wide range of maternal endothelial dysfunction [20]. It was reported that hucMSC-Exo improved the morphology of placental tissue in PE rats through suppressing cell apoptosis and facilitating angiogenesis in placental tissue in a dose-dependent manner [8]. A study has reported that miR-18b expression affected cell migration, viability and invasion in PE [12]. Moreover, it was verified increased maternal LEP concentration and hypomethylation of the LEP in placenta in early onset PE [21]. The current study was designed to explore the effect of exosomes and miR-18b-3p targeted LEP on the occurrence of PE. The findings in this study revealed that hucMSC-derived exosomal miR-18b-3p inhibited PE progression by reducing LEP.
Based on our findings, miR-18b-3p reduced and LEP elevated in placenta tissues of PE rats. Similar to our study, the mRNA expression of miR-18b was markedly suppressed in PE placental tissues relative to that in normal placental tissues [12]. In addition, a study revealed that miR-18b content was dramatically reduced in malignant melanoma tissues in comparison with their matched adjacent non-tumor tissues [22]. Another study has verified that placental LEP expression was raised in preterm PE compared with controls [23]. Moreover, a study showed that LEP expression was obviously heightened in preeclamptic placentas [15]. This literature provided a theoretical basis for us to explore the abnormal expression of miR-18b-3p and LEP in PE. Moreover, it was predicted using a bioinformatic software that LEP was targeted by miR-18b-3p, and this targeting relationship was further confirmed with dual-luciferase reporter gene assay in our research. A study reported that LEP is a target for all three miRNAs (miR-1301, miR-223 and miR-224) in early-onset PE [16]. Another study has displayed that LEP decreased miR-93 expression in osteoarthritis and rheumatoid arthritis [24]. However, the binding between miR-18b-3p and LEP in human diseases, especially in PE, remains scarcely studied, which is the novelty of this study. Furthermore, a result emerging from our study reported that exosomes increased miR-18b-3p and decreased LEP in placenta tissues of PE. It was formerly documented that the expression of miR-18b-5p was notably raised in colorectal cancer plasma exosomes [25], while the relationship between hucMSC-Exos and miR-18b-3p/LEP in PE needs further study.
Additionally, the finding from our investigation showed that restored miR-18b-3p reduced SBP and 24-h proteinuria of PE rats, increased the weight of placenta, declined TNF-α, IL-1β and IL-6 contents in serum and placenta tissues as well as suppressed cell apoptosis. These data indicated that miR-18b-3p elevation contributes to alleviating the symptoms and pathological changes in PE. It was demonstrated that stable upregulation of miR-18b produced effective tumor inhibitor activity, such as inhibiting melanoma cell viability, inducing apoptosis and reducing tumor growth in vivo [26]. Another result in our study was that hucMSC-Exos reduced SBP and 24-h proteinuria of PE rats, increased the weight of placenta, declined TNF-α, IL-1β and IL-6 contents in serum and placenta tissues as well as suppressed cell apoptosis. The findings of the current study revealed that exosomes treated PE rat models presented an increase of the number and quality of fetuses, the quality of placenta, but cell apoptosis was significantly reduced [8]. Interestingly, a previous research has demonstrated that the addition of fetal bovine exosomes declined contents of macrophage TNF-α and IL-6 [27]. A study has revealed that purified exosomes suppressed production of IL-1β in lipopolysaccharide/nigericin-stimulated macrophages [28]. Furthermore, Nong et al. have suggested that inflammatory markers, such as TNF-α and IL-6, were dramatically decreased after administration of exosomes produced through human-induced pluripotent stem cell-derived MSCs [29]. There is a article finding that the SBP was markedly elevated in the group of women who later developed PE [30, 31]. It was displayed that PE patients were positively associated with SBP and diastolic blood pressures and proteinuria [32]. Also, a recent study has provided a proof that proteinuria heightened with advancing gestation in PE women [33]. A important finding was that rats from the PE group had increased TNF-α relative to the normal pregnant group [34]. Another study has verified that serum IL-6 and IL-1β were obviously elevated in women with PE in relation to controls [35]. The above findings suggested that PE patients usually showed high SBP, proteinuria and levels of inflammatory factors. Thus, it could be inferred from our results that the hucMSC-derived exosomal miR-18b-3p had a therapeutic effect on PE.
Conclusion
In conclusion, our study provides evidence that hucMSCs-derived exosomes upregulate miR-18b-3p, which targets LEP to suppress the contents of inflammatory factors and reduce cell apoptosis rate in PE rat placenta tissues, thereby inhibiting the occurrence of PE. Thus, exosomal miR-18b-3p may be a potential candidate for treatment of PE via targeting LEP. This research identified the role of hucMSC-derived exosomal miR-18b-3p targeting LEP during PE development for the first time, which provided a novel insight for PE treatment. However, due to the limitation of known researches, the study needs to be monitored rigorously and reported appropriately in the future clinical trials.
データと資料の可用性
該当なし。
ナノマテリアル