


microRNA-499-5pの復元は、Sox6を標的とすることで敗血症誘発性肺損傷マウスを保護します
要約
背景
マイクロRNA(miR)は敗血症に関与することが知られています。したがって、我々は、マウスの敗血症誘発性肺損傷に対する性決定領域Y関連の高移動度群ボックス6(Sox6)を標的とするmiR-499-5pの保護効果について議論することを目指しています。
メソッド
敗血症誘発性肺損傷モデルは、盲腸結紮および穿刺によって確立された。マウスの肺組織における湿重量/乾重量(W / D)比、miR-499-5p、Sox6、カスパーゼ-3、およびカスパーゼ-9の発現をテストしました。肺損傷スコア、コラーゲン線維、および肺組織における肺線維症の程度が決定された。さらに、肺組織の細胞アポトーシスを測定した。気管支肺胞洗浄液(BALF)と肺組織の炎症性因子の含有量と酸化ストレス指数は、機能喪失および機能獲得アッセイによって検出されました。 miR-499-5pとSox6のターゲティング関係を確認しました。
結果
敗血症誘発性肺損傷マウスの肺組織では、W / D比とSox6が増加し、miR-499-5pが減少しました。 miR-499-5pの回復またはSox6の枯渇により、肺組織の病理が緩和され、肺損傷スコア、コラーゲン線維、肺線維症の程度、TUNEL陽性細胞、カスパーゼ3およびカスパーゼ9タンパク質の発現、およびBALFおよび肺組織における炎症性因子の含有量が減少しました。敗血症誘発性肺損傷マウスの肺組織における酸化ストレス応答と同様に。 miR-499-5pはSox6をターゲットにしました。
結論
miR-499-5pの高発現は、肺組織の細胞アポトーシスを弱め、Sox6の枯渇を介して敗血症誘発性肺損傷マウスの炎症を抑制することができ、敗血症誘発性肺損傷の潜在的な候補マーカーおよび治療標的です。
はじめに
敗血症は、感染の結果として発生し、急性呼吸窮迫症候群を含む複数の臓器機能障害を引き起こす全身性炎症です[1]。それは、重度の低血圧、進行性低酸素血症、組織損傷および炎症性変化を特徴とします[2]。肺は敗血症で最も重要で脆弱な臓器であり、急性肺損傷(ALI)は頻繁な敗血症誘発性炎症性疾患です[3]。敗血症によって誘発されるALIの重症度は、肺胞毛細血管網の透過性の障害に起因する可能性があり、その結果、酸素供給が低下します[4]。肺細胞のアポトーシスは、敗血症によって誘発される肺損傷の重要な要因です[5]。敗血症誘発性ALIの病態生理学は不明であり、敗血症およびALI患者で利用できる治療法は支持療法と抗生物質のみであり、敗血症による高い死亡率への影響は限定的です[6]。
マイクロRNA(miRNA)は、内因的に発現するRNAの小さな非コードセグメントであり、約18〜24ヌクレオチドで構成され、広範囲の細胞機能を制御し、特定の遺伝子の標的領域に結合して、mRNAの阻害または活性化を通じて発現を制御できます。翻訳/転写[7]。 MiR-499-5pは、最近発見されたミオシンでエンコードされたmiRNAのメンバーです[8]。ある研究では、miR-499-5pの血清レベルを利用して敗血症患者の臓器不全を示すことができると報告されています[9]。別の研究では、miR-499-5pの発現が敗血症によって誘発される心臓のアポトーシスに関与していることが明らかになっています[10]。性決定領域Y(SRY)関連の高移動度ボックス(HMG)(SOX)ファミリーは、SRYとの配列同一性が50%を超えるHMGクラスのDNA結合ドメインを含む一連の遺伝子によって定義されます[11]。 。 Sox6転写因子はSRY関連のHMGボックスファミリーの一部であり、進行中に複数の組織で発現し、増殖中の前駆細胞から機能的に成熟した細胞への移行に重要な役割を果たします[12]。上昇したSox6が敗血症誘発性心臓アポトーシスに関係していることを報告している研究があります[10]。研究によると、Sox2の発現とSox4の発現はヒトの小細胞肺がんで増強されることが示されています[13、14]。しかし、Sox6と敗血症による肺損傷との関係については、さらに研究が必要です。したがって、私たちの研究は、Sox6を標的とすることにより、敗血症誘発性肺損傷マウスに対するmiR-499-5pの保護効果を調査することでした。
材料と方法
倫理基準への準拠
すべての動物実験は、国立衛生研究所の実験動物の管理と使用に関するガイドに沿ったものでした。この議定書は、山東大学チールー医科大学威海市立病院の動物実験倫理委員会によって許可されました。
実験動物
健康なオスの特定病原体除去グレードのBABL / cマウス(6〜8週齢、体重18〜22 g)は、山東大学(中国山東省)の実験動物センターから購入しました。環境は24±2°Cに設定され、12の明暗サイクルが交互に繰り返されました。マウスは、無菌食品および滅菌水を自由に摂取できる状態で処理された。実験は7日間の適応給餌後に開始されました。
マウスの盲腸結紮穿刺(CLP)による敗血症誘発性肺損傷モデルの確立
マウスはランダムに2つのグループに分けられました:偽のグループ( n =8)およびモデルグループ。手術前にマウスを12時間絶食させた後、腹腔内注射により1%ペントバルビタールナトリウム(70 mg / kg)で麻酔をかけました。麻酔の効果が観察されました。マウスの呼吸と心拍に気づいた。マウスの頭と手足を仰臥位に固定し、腹部の皮膚を準備し、ヨードフォアコットンボールで消毒した。モデル群のマウスは、腹部正中線の中央部分を縦切開(1.0cm)して治療し、腹腔を開き、腸間膜血管の損傷を避けて盲腸を腹腔から引き抜いた。盲腸の端の1/2で1–0の絹糸で結紮し、盲腸を腸間膜の反対側の縁に穿刺針No.16で穿刺しました。適切な量の腸内容物を押し出しました。盲腸を元に戻し、切開部を縫合した。腹腔を開いた後、偽群のマウスは盲腸のみを引き抜いた後、盲腸を元に戻した。手術後、マウスに0.9%塩化ナトリウム溶液1 mLを皮下注射し、ケージに入れて定期的に給餌しました。
インビボトランスフェクション
モデリングの前に、マウスは7つのグループに割り当てられました( n =8):モデルグループ、miR-499-5pアゴミールグループ、アゴミールネガティブコントロール(NC)グループ、低分子干渉RNA(si)-Sox6グループ、si-NCグループ、miR-499-5pアゴミール+過剰発現(oe)-Sox6グループおよびmiR-499-5pアゴミール+ oe-NCグループ。モデリングの1時間前に、グループ化に従ってマウスを治療しました。マウスを腹腔内注射により1%ペントバルビタールナトリウムで麻酔し、次に45°の傾斜の仰臥位で固定し、マウスの舌を口の反対側に動かして気道の開通性を確保した。マウスの首の前部皮膚を消毒し、気管を露出させるために切断した。トランスフェクション複合体(50μL)をマイクロサンプラーで吸収し(グループに応じて滴下し、同量のリン酸緩衝生理食塩水(PBS)をモデルグループに滴下しました)、ゆっくりと気管に滴下しました。トランスフェクション後、マウスを垂直に回転させ、トランスフェクション複合体を肺に完全に分配しました[15]。トランスフェクションプロセス中に、マウスの呼吸と心拍が観察されました。呼吸停止と心停止が発生した場合、トランスフェクションは直ちに停止され、気道は遮られませんでした。マウスの呼吸と心拍が安定したときにトランスフェクションを続けた。 MiR-499-5pアゴミールとそのNC、および対応するSox6のプラスミドは、Shanghai GenePharma Co.、Ltd。(Shanghai、China)から購入しました。トランスフェクション後、マウスをケージで分離し、1日間飼育し、安楽死させました。マウスを腹腔内注射により1%ペントバルビタールナトリウムで麻酔し、仰臥位に固定し、腹部大動脈静脈切開により安楽死させた。マウスの皮膚を局所的に消毒し、胸腔を開き、気管と右主気管支を結紮し、右主気管支を切り離し、右肺を摘出した。病理検査のために右肺の下葉を10%中性ホルマリンに入れ、中葉をすばやく秤量して湿重量(W)を計算し、56℃で焼き、72時間後に乾燥重量(D)を計算しました。したがって、W / D比が計算されました。右上葉は、mRNAおよびタンパク質の検出のために液体窒素に保存されました。左肺を0.4mLの生理食塩水で3回洗浄し、約1 mLの気管支肺胞洗浄液(BALF)を収集し、1.5 mLのエッペンドルフ(EP)チューブに移して、氷上に置きました。 BALFを1500rpmで5分間遠心分離し、新しいEPチューブに移してから、炎症性因子を検出するために-80℃で保存しました。
さらに、マウスを3つのグループに分けました( n =10 /グループ):偽のグループ、モデルグループ、およびmiR-499-5pアゴミールグループ(50μLのmiR-499-5pアゴミールを手術の1時間前に気管に注入しました)。生存率は12時間ごとに、完全に7日間監視されました。 7日以内のマウスの生存状態が観察され、miR-499-5pの毒性が決定されました[16]。
ヘマトキシリン-エオシン(H&E)染色および損傷スコア
肺組織をパラフィン切片に調製し、H&Eで染色し、光学顕微鏡で観察しました。肺損傷は、米国胸部学会によって発行された肺損傷の病理学的スコアリングシステムによってスコアリングされました(肺胞うっ血、出血、好中球浸潤または肺胞腔または血管壁の凝集、肺胞壁肥厚および/または硝子膜形成)。 0ポイントは病変がないか非常に軽度であることを示しました。 1点は軽度の病変を示した。 2点は中等度の病変を示した。 3点は重度の病変を示した。 4点は非常に重度の病変を示した。 4つの項目の合計スコアは、肺損傷の合計スコアを表しています[17]。
マッソン染色とスコア
肺組織をパラフィン切片(4 µm)にし、マッソン染色で処理しました[18]。コラーゲン線維、粘液、軟骨は青、細胞質、筋肉、セルロース、グリアは赤、細胞核は青黒でした。半定量的アシュクロフトスコアリングシステム[19]では、0ポイントは正常な肺を示しました。 1点は、肺胞または気管支壁の軽度の線維性肥厚を示した。 3点は、肺構造に損傷を与えることなく、肺胞または細気管支壁が中程度に肥厚していることを示しています。 5点は、肺構造の損傷を伴う線維症の悪化、繊維バンドまたは腫瘤の形成を示しました。 7点は、重度の肺構造の損傷と線維症の広い領域を示し、繊維バンドまたは凝集塊を形成しました。 8ポイントは、すべての地域の繊維状パッキングを示しています。
末端デオキシヌクレオチドトランスフェラーゼを介したデオキシウリジン三リン酸-ビオチンニックエンドラベリング(TUNEL)染色
肺組織のアポトーシスは、TUNELキット(Roche Diagnostics GmbH、マンハイム、ドイツ)によってテストされ、光学顕微鏡によって観察されました。アポトーシス細胞は核内で茶色に染色された。アポトーシス指数は、TUNEL陽性細胞の割合を示しました[20]。
免疫組織化学
肺組織切片をキシレンで脱パラフィンし、3%過酸化水素でインキュベートし、ヤギ血清でブロックしました。次に、カスパーゼ-3(ab13847、1:500)、カスパーゼ-9(ab32539、1:250)、およびSOX6(ab30455、1:500、すべて米国マサチューセッツ州アブカム)を免疫染色切片の抗体として使用しました。光学顕微鏡(Leica Microsystems、IL、USA)を使用して画像をキャプチャし、Image Pro Plus 6.0ソフトウェア(Media Cybernetics、Rockville、MA、USA)で分析しました[21]。
逆転写定量的ポリメラーゼ連鎖反応(RT-qPCR)
肺組織中のトータルRNAの抽出とRNA濃度の測定には、OMEGAキット(Solarbio Science&Technology Co.、Ltd。、北京、中国)を採用しました。相補的DNA(cDNA)は、RevertAid First Strand cDNA合成キット(Thermo Fisher Scientific、マサチューセッツ、米国)に沿った逆転写によって構成されました。プライマーは、表1に示すように、Invitrogen(Carlsbad、CA、USA)によって設計されました。PCR反応は、FastStart Universal SYBR Green Master(Rox)キット(Roche)に従って実行されました。実験は2 -△△ Ct によって定量的に分析されました 方法。 U6はmiR-499-5pのローディングコントロールであり、GAPDHは他の遺伝子の内部パラメーターでした[22]。
<図>ウエスタンブロット分析
肺組織を治療するために、放射性免疫沈降アッセイ溶解緩衝液を使用して総タンパク質を収集した。総タンパク質濃度は、ビシンコニン酸法によって測定されました。タンパク質サンプル(30μg)を10%ドデシル硫酸ナトリウムポリアクリルアミドゲル電気泳動で処理し、ポリビニリデンフルオリドメンブレンに転写し、5%スキムミルクパウダーでブロックし、一次抗体Sox6(ab30455、1:1000、Abcam)、カスパーゼと反応させました。 -3(9661、1:1000、Cell Signaling Technology、MA、USA)、Caspase-9(9502、1:1000、Cell Signaling Technology)、腫瘍壊死因子-α(TNF-α、ab6671、1:1000、Abcam )、インターロイキン-1β(IL-1β、16806-1-AP、1:500、Proteintech、USA)、IL-6(ab6672、1:1000、Abcam)およびGAPDH(sc-32233、1:1000、Santa Cruz Biotechnology、Inc.、CA、USA)。次に、メンブレンを二次抗体(1:2000)とインキュベートし、化学発光を増強して発色させました[23]。
酵素結合免疫吸着測定法(ELISA)
BALF中のIL-1β、IL-6およびTNF-αのレベルは、ELISAキット(Boster、湖北省、中国)によって決定されました。比色定量は450nmで行いました。
酸化ストレス指数の検出
液体窒素中の肺組織を秤量し、低温条件下で一定の割合の生理食塩水で希釈した後、均質化し、低温で4℃、3000 r / minで20分間遠心分離し、上清を採取して測定しました。ミエロペルオキシダーゼ(MPO)の活性は、MPO検出キット(南京建成生物工学研究所、南京、中国)によってテストされ、マロンジアルデヒド(MDA)、スーパーオキシドジスムターゼ(SOD)、グルタチオンペルオキシダーゼ(GSH-Px)およびカタラーゼ(CAT)がテストされました。 JianCheng BioengineeringInstituteが提供するキットによる。
デュアルルシフェラーゼレポーター遺伝子アッセイ
Sox6は、生物学的Webサイト(http://www.targetscan.org/vert_72/)によって予測されたmiR-499-5pのターゲット遺伝子である可能性があります。 miR-499-5pとSox6mRNA 3 '非翻訳領域(UTR)の間には4つの結合部位がありました。 pmiR-RB-REPORT-Sox6野生型(WT)およびpmiR-RB-REPORT-Sox6変異型(MUT)ベクターは、Guangzhou RiboBio Co.、Ltd。(広東省、中国)によって構築され、対応するルシフェラーゼレポータープラスミドは生成されます。 TC-1細胞(マウス肺上皮細胞、1×10 5 細胞/ウェル、Shanghai Yiyan biotechnology Co.、Ltd.、Shanghai、China)を24ウェルプレートに播種し、miR-499-5pアゴミールまたはアゴミール-NC、pmiR-RB-REPORT-Sox6WTまたはpmiR-RB-REPORT-Lipofectamine2000を介したSox6MUT。デュアルルシフェラーゼレポーター遺伝子アッセイシステム(Promega、WI、USA)を適用して、ルシフェラーゼ活性を測定しました。
統計分析
すべてのデータは、SPSS 21.0(IBM Corp. Armonk、NY、USA)ソフトウェアによって分析されました。測定データは、平均±標準偏差で伝達されました。 2つのグループ間の比較は、 t によって行われました。 テストでは、複数のグループ間の比較は一元配置分散分析(ANOVA)によって評価され、Tukeyの事後テストはANOVA分析後のペアワイズ比較に使用されました。生存曲線はカプランマイヤー法で作成されました。 P 値<0.05は統計的に有意な差を示しました。
結果
敗血症誘発性肺損傷マウスの肺組織ではmiR-499-5pが減少する一方で、W / D比とSox6が増加します
miR-499-5pの毒性をマウスでテストして、敗血症マウスの生存期間を検出しました。結果は(図1a)、偽のグループのマウスの7日以内の生存率が100%であり、敗血症のマウスの生存率が低下したことを示しました。 miR-499-5p治療後の敗血症マウスの生存率が改善されました。肺組織のW / D比の結果は、(図1b):偽のグループと比較して、モデルグループ( P )でW / D比が上昇したことを示しました。 <0.05)。アゴミールNCグループとsi-NCグループと比較して、miR-499-5pアゴミールグループとsi-Sox6グループ(両方 P )でW / D比が低下しました。 <0.05)。 W / D比は、miR-499-5pアゴミール+ oe-NCグループと比較してmiR-499-5pアゴミール+ oe-Sox6グループで著しく高くなりました( P <0.05)。
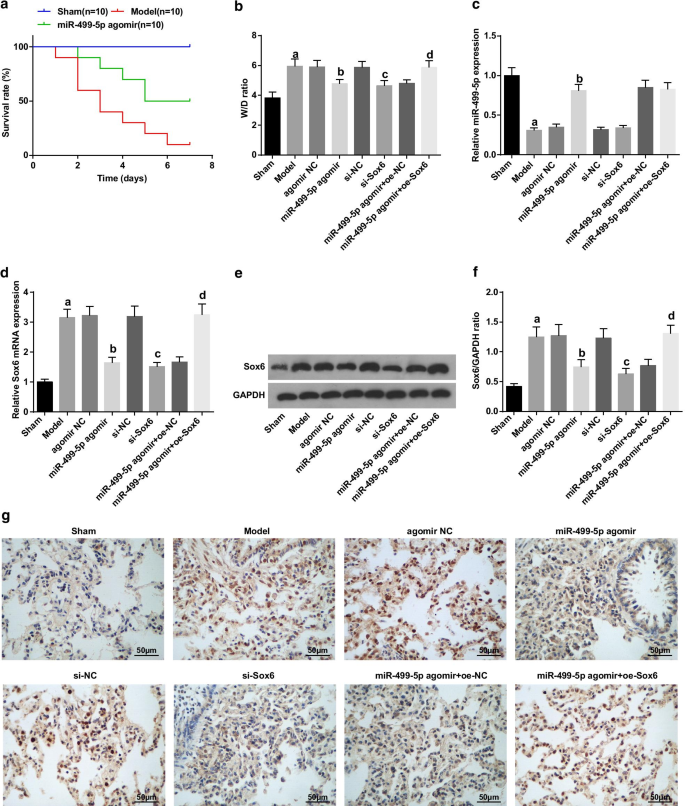
敗血症誘発性肺損傷マウスの肺組織では、W / D比とSox6が増加し、miR-499-5pが減少します。 a Kaplan–Meierによって作成されたマウスの生存曲線。 b 各グループの肺組織のW / D比。 c マウスの肺組織におけるmiR-499-5pの発現。 d マウスの肺組織におけるSox6mRNAの発現。 e ウエスタンブロット分析によるSox6のタンパク質バンド。 f マウスの肺組織におけるSox6のタンパク質発現。 g マウスの肺組織のSox6免疫組織化学。 (a) P <0.05対偽のグループ。 (b) P <0.05対アゴミールNCグループ。 (c) P <0.05対si-NCグループ。 (d) P <0.05対miR-499-5pアゴミール+ oe-NCグループ。測定データは平均±標準偏差として表され、データは一元配置分散分析とそれに続くテューキーの事後検定によって評価されました
MiR-499-5pとSox6の発現が検出され、次のことがわかりました(図1c–f):偽のグループとは対照的に、モデルグループではmiR-499-5pが減少し、Sox6が上昇しました(両方 P <0.05)。アゴミールNCグループに関連して、miR-499-5pアゴミールグループではmiR-499-5pが上昇し、Sox6が減少しました(両方 P <0.05)。 si-NCグループと比較して、Sox6はsi-Sox6グループで落ち込んでいます( P <0.05)。 miR-499-5pアゴミール+ oe-NCグループと比較すると、Sox6はmiR-499-5pアゴミール+ oe-Sox6グループで劇的に上昇しました( P <0.05)。
敗血症マウスの肺組織におけるSox6のタンパク質含有量は、免疫組織化学によって決定されました(図1g)。偽のグループと比較して、Sox6タンパク質の発現がモデルグループで増加したことが観察されました( P <0.05);アゴミールNCグループおよびsi-NCグループと比較して、miR-499-5pアゴミールグループおよびsi-Sox6グループ(両方 P )でより低いSox6タンパク質発現が調べられました。 <0.05); miR-499-5pアゴミール+ oe-NCグループと比較して、Sox6タンパク質の発現はmiR-499-5pアゴミール+ oe-Sox6グループで増強されました( P <0.05)。
復元されたmiR-499-5pまたは枯渇したSox6は、肺組織の病状を緩和し、肺損傷スコア、コラーゲン線維、および敗血症誘発性肺損傷における肺線維症の程度を低下させますマウス
HE染色の結果は、(図2a):偽のグループでは、肺組織構造は無傷であり、肺胞中隔は基本的に浮腫や炎症なしに現れ、肺胞腔は透明であったことを示しました。モデルでは、agomir NC、si-NC、miR-499-5p agomir + oe-Sox6グループ、滲出、浮腫、出血が肺組織のほとんどの肺胞腔に見られ、炎症細胞の大量浸潤が肺間質に見られました。 miR-499-5pアゴミール、si-Sox6、miR-499-5pアゴミール+ oe-NCグループでは、モデルグループと比較して肺組織の病変が減少しました。
復元されたmiR-499-5pまたは枯渇したSox6は、肺組織の病理を緩和し、敗血症誘発性肺損傷マウスの肺損傷スコア、コラーゲン線維、および肺線維症の程度を低下させます。 a マウスの肺組織におけるHE染色の結果。 b 各グループのマウスの肺組織損傷スコア。 c マウスの肺組織のマッソン染色。 d 各グループのマウスの肺線維症スコア。 (a) P <0.05対偽のグループ。 (b) P <0.05対アゴミールNCグループ。 (c) P <0.05対si-NCグループ。 (d) P <0.05対miR-499-5pアゴミール+ oe-NCグループ。測定データは平均±標準偏差として表され、データは一元配置分散分析とそれに続くテューキーの事後検定によって評価されました
マウスの肺損傷スコアと半定量的アシュクロフトスコアの結果は、次のことを示しました(図2b、d):偽のグループに関連して、肺損傷スコアと肺線維症の程度はモデルグループで明らかに増強されました( P > <0.05)。アゴミールNCグループおよびsi-NCグループと比較して、肺損傷スコアおよび肺線維症の程度は、miR-499-5pアゴミールおよびsi-Sox6グループ(両方 P )で減少しました。 <0.05)。 miR-499-5pアゴミール+ oe-NCグループとは対照的に、miR-499-5pアゴミール+ oe-Sox6グループでは肺損傷スコアと肺線維症の程度が著しく上昇しました( P <0.05)。
マッソン染色の結果は、(図2c):偽のグループでは、マウスの肺組織に少量の青いコラーゲン線維が含まれていることを明らかにしました。モデル、アゴミールNC、si-NCおよびmiR-499-5pアゴミール+ oe-Sox6グループでは、コラーゲン線維が増加しました。 miR-499-5pアゴミールグループ、si-Sox6グループ、miR-499-5pアゴミール+ oe-NCグループでは、モデルグループと比較してコラーゲン線維が減少しました。
復元されたmiR-499-5pまたは枯渇したSox6は、敗血症誘発性肺損傷マウスの肺組織におけるTUNEL陽性細胞、カスパーゼ-3およびカスパーゼ-9タンパク質の発現を低下させます
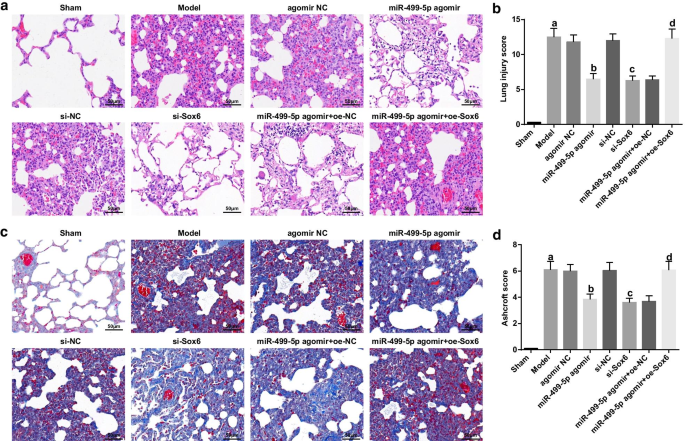
ウエスタンブロット、TUNEL染色、および免疫組織化学の結果は、(図3a–f):正常な核は青色で、アポトーシス核はさまざまな色合いで茶色であることを示しました。偽のグループと比較して、TUNEL陽性細胞、カスパーゼ-3およびカスパーゼ-9タンパク質の発現はモデルグループで強化されました(すべて P <0.05)。アゴミールNCおよびsi-NCグループと比較して、TUNEL陽性細胞、カスパーゼ-3およびカスパーゼ-9タンパク質の発現は、miR-499-5pアゴミールおよびsi-Sox6グループ(すべて P )で減少しました。 <0.05)。 miR-499-5pアゴミール+ oe-NCグループと比較して、TUNEL陽性細胞、カスパーゼ-3およびカスパーゼ-9タンパク質の発現は、miR-499-5pアゴミール+ oe-Sox6グループ(すべて P <0.05)。
復元されたmiR-499-5pまたは枯渇したSox6は、敗血症誘発性肺損傷マウスの肺組織におけるTUNEL陽性細胞、カスパーゼ-3およびカスパーゼ-9タンパク質の発現を低下させます。 a マウスの肺組織におけるTUNEL陽性細胞。 b マウスの肺組織におけるTUNEL陽性細胞の数。 c マウスの肺組織におけるカスパーゼ-3およびカスパーゼ-9の免疫組織化学; d マウスの肺組織におけるカスパーゼ-3およびカスパーゼ-9の平均OD値。 e マウスの肺組織におけるカスパーゼ-3およびカスパーゼ-9のタンパク質バンド。 f 各グループのマウスの肺組織におけるカスパーゼ-3およびカスパーゼ-9タンパク質の発現。 (a) P <0.05対偽のグループ。 (b) P <0.05対アゴミールNCグループ。 (c) P <0.05対si-NCグループ。 (d) P <0.05対miR-499-5pアゴミール+ oe-NCグループ。測定データは平均±標準偏差として表され、データは一元配置分散分析とそれに続くテューキーの事後検定によって評価されました
復元されたmiR-499-5pまたは枯渇したSox6は、敗血症誘発性肺損傷のBALFおよび肺組織のTNF-α、IL-1β、およびIL-6含有量を減少させますマウス
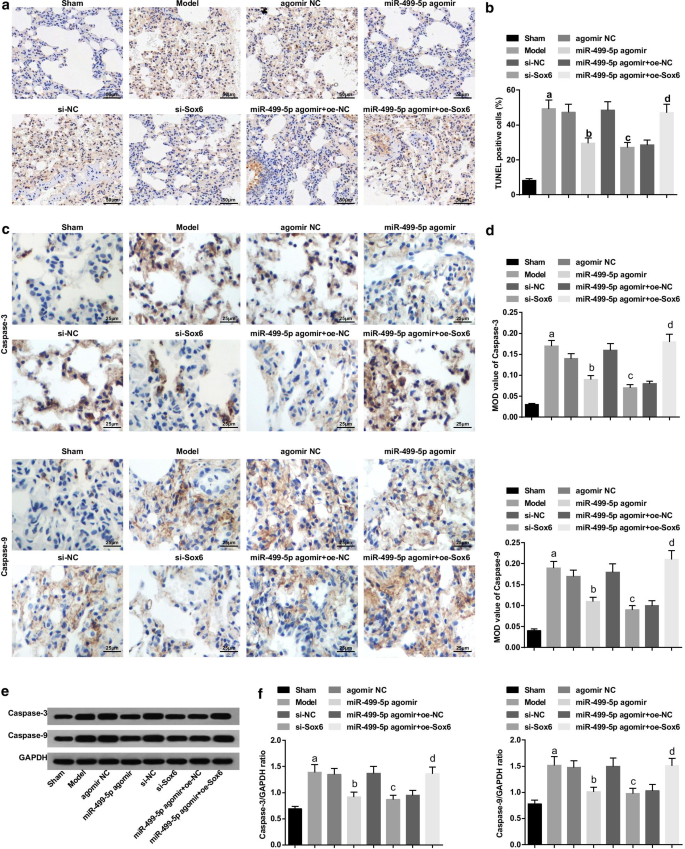
ELISAおよびウエスタンブロットアッセイの結果により、(図4a–g):TNF-α、IL-1βおよびIL-6の含有量が偽のグループと比較してモデルグループで増加したことが示されました(すべて P <0.05)。アゴミールNCおよびsi-NCグループと比較して、TNF-α、IL-1βおよびIL-6の含有量は、miR-499-5pアゴミールおよびsi-Sox6グループ(すべて P )で減少しました。 <0.05)。 miR-499-5pアゴミール+ oe-NCグループと比較して、TNF-α、IL-1βおよびIL-6の含有量はmiR-499-5pアゴミール+ oe-Sox6グループ(すべて P > <0.05)。
回復したmiR-499-5pまたは枯渇したSox6は、敗血症誘発性肺損傷マウスのBALFおよび肺組織におけるTNF-α、IL-1β、およびIL-6の含有量を減少させます。 a 各グループのマウスのBALFにおけるTNF-α含有量。 b 各グループのマウスのBALFにおけるIL-1β含有量。 c 各グループのマウスのBALFにおけるIL-6含有量。 d 各グループのマウスの肺組織におけるTNF-α、IL-1βおよびIL-6のタンパク質バンド。 e 各グループのマウスの肺組織におけるTNF-αのタンパク質発現。 f 各グループのマウスの肺組織におけるIL-1βのタンパク質発現。 G 各グループのマウスの肺組織におけるIL-6のタンパク質発現。 (a) P <0.05対偽のグループ。 (b) P <0.05対アゴミールNCグループ。 (c) P <0.05対si-NCグループ。 (d) P <0.05対miR-499-5pアゴミール+ oe-NCグループ。測定データは平均±標準偏差として表され、データは一元配置分散分析とそれに続くテューキーの事後検定によって評価されました
復元されたmiR-499-5pまたは枯渇したSox6は、SOD、CAT、およびGSH-Pxの活動を高め、敗血症によって誘発された肺組織のMDAおよびMPOの含有量を減らします肺損傷マウス
酸化ストレス指数の検出結果は、(図5a–e):偽のグループに対して、モデルグループでMDAおよびMPOの含有量が増加する一方で、SOD、CAT、およびGSH-Pxの活動が損なわれることを示しました(すべて P > <0.05)。アゴミールNCおよびsi-NCグループと比較して、SOD、CAT、およびGSH-Pxの活動は増加し、MDAおよびMPOの含有量はmiR-499-5pアゴミールおよびsi-Sox6グループ(すべて P )で減少しました。 <0.05)。 miR-499-5pアゴミール+ oe-NCグループと比較して、SOD、CAT、GSH-Pxの活動は抑制され、MDAとMPOの内容はmiR-499-5pアゴミール+ oe-Sox6グループ(すべて P <0.05)。
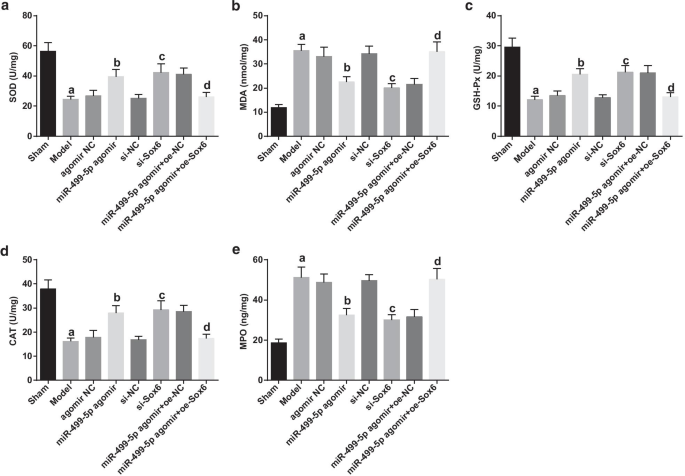
復元されたmiR-499-5pまたは枯渇したSox6は、SOD、CAT、およびGSH-Pxの活動を高め、敗血症誘発性肺損傷マウスの肺組織におけるMDAおよびMPOの含有量を減らします。 a マウスの肺組織におけるSOD活性。 b マウスの肺組織におけるMDA含有量。 c マウスの肺組織におけるGSH-Px活性。 d マウスの各グループの肺組織におけるCAT活性。 e マウスの肺組織のMPO含有量。 (a) P <0.05対偽のグループ。 (b) P <0.05対アゴミールNCグループ。 (c) P <0.05対si-NCグループ。 (d) P <0.05対miR-499-5pアゴミール+ oe-NCグループ。測定データは平均±標準偏差として表され、データは一元配置分散分析とそれに続くテューキーの事後検定によって評価されました
miR-499-5pはSox6を直接ターゲットにします
バイオインフォマティクスのWebサイトTargetScanを通じて、miR-499-5pとSOX4 / 6の間に標的結合部位があると予測しました。 RT-qPCRを使用すると、偽のグループと比較して、モデルグループでSOX2 / 4/6の発現が増加し、SOX6の発現がほとんど増加することがわかりました(すべて P <0.05);アゴミールNCグループと比較して、miR-499-5pアゴミールグループはSOX2発現の変化を示さなかったが、SOX4発現の低下、主にSOX6の低下を示した(追加ファイル1:図S1a、b)。そのため、最終的にmiR-499-5pの標的遺伝子としてSOX6を選択しました。生物学的予測のウェブサイト(http://www.targetscan.org/vert_72/)は、miR-499-5pがSox6を標的にする可能性があることを明らかにしました(図6a)。デュアルルシフェラーゼレポーター遺伝子アッセイは、対照群と比較して、pmiR-RB-REPORT-Sox6 WTベクターとmiR-499-5pアゴミールをTC-1細胞にコトランスフェクトした後、ルシフェラーゼ活性が明らかに破壊されたことを示しました( P <0.05)、ルシフェラーゼ活性は、miR-499-5pアゴミールをpmiR-RB-REPORT-Sox6 MUTベクターでコトランスフェクトした後、またはアゴミール-NCをpmiR-RB-REPORT-Sox6 WTベクターでコトランスフェクトした後、明確な違いを示しませんでした( P > 0.05)、miR-499-5pがSox6 3'UTR WTに直接結合し、ルシフェラーゼ活性を阻害する可能性があることを示唆しています。

miR-499-5pはSox6を直接ターゲットにしています。 a 生物学的ウェブサイトによって予測されるmiR-499-5pとSox6の間のターゲット関係。 b デュアルルシフェラーゼレポーター遺伝子アッセイの検証結果。 * P <0.05対模倣NCグループ。測定データは平均±標準偏差として表され、2つのグループ間の比較は t によって行われました。 テスト
ディスカッション
敗血症はショックおよび多臓器不全症候群の重大な原因であり、その中でALIは最も頻繁で重大な臓器合併症です[24]。敗血症誘発性ALIは、最も早く、最も高い発生率であるだけでなく、急速に進行し、致死率が高く、現在、効果的な治療法はありません[25]。以前の研究では、miR-218のアップレギュレーションが炎症性因子の分泌を弱め、敗血症による肺損傷を予防したとされています[26]。また、miR-146aのレベルを促進すると、ALIの炎症を軽減でき[27]、miR-125bの回復により、ALIのマウスの生存率が向上し、炎症が抑制されます[28]。研究は肺損傷におけるmiRNAの役割を強調していますが、敗血症誘発性肺損傷におけるmiR-499-5pの詳細なメカニズムはほとんど調査されていないため、敗血症誘発性におけるその治療可能性を発見するためにmiR-499-5pに焦点を当てました。 ALI。また、ある研究では、Sox6の増加が敗血症によって誘発される心臓のアポトーシスに関与しているという証拠が提供されています[10]。敗血症による肺損傷を深く理解するための努力がなされるべきである。敗血症誘発性肺損傷マウスに対するSox6を標的とするmiR-499-5pの保護効果について議論することを目指しています。
我々の研究では肺組織のW / D比を計算し、マウス肺組織におけるSox6とmiR-499-5pの発現を評価しました。結果は、敗血症誘発性肺損傷マウスの肺組織において、miR-499-5pが減少する一方で、W / D比およびSox6が増加することを示した。 A recent study has proposed that miR-499-5p expression in patients with mild sepsis, severe sepsis and septic shock was dramatically decreased relative that in normal controls [9]. Another study has presented that miR-499-5p expression was markedly declined in non-small cell lung cancer (NSCLC) tissues and linked to poor clinical outcomes [29]. It is reported that the Sox6 expression was raised in lung cancer tissues in comparison with normal tissues [30]. Similarly, a previous study has revealed that the expression of Sox6 was remarkably elevated in the kidney tissues of mice with diabetic kidney disease [31]. Our study also presented that miR-499-5p directly targeted to Sox6. An important finding was that Sox6 is a target gene of miR-499 [10]. Another study provided data of that Sox6 was a target gene of miR-499-5p, and restoration of miR-499-5p suppressed the expression of Sox6 [32]. However, the target relation of miR-499-5p and Sox6 in lung tissues of sepsis-induced mice has not been uncovered yet.
In addition, it was revealed that restored miR-499-5p or depleted Sox6 alleviated lung tissues pathology, reduced lung injury score, collagen fibers and the degree of pulmonary fibrosis, and reduced TUNEL positive cells, Caspase-3 and Caspase-9 protein expression in lung tissues of sepsis-induced lung injury mice. It has been previously suggested that up-regulating miR-499-5p suppresses cell proliferation and promotes apoptosis in vivo and in vitro of NSCLC [29]. Another study has verified that restoring miR-499-5p and depleting Sox6 attenuated hypoxia/re-oxygenation-induced cell apoptosis and reduced Caspase-3 expression [32]. It was presented that down-regulating Sox6 alleviates the pathological injury and represses neuronal apoptosis in hippocampal tissues of Alzheimer’s disease [33]. Moreover, a study revealed that depletion of Sox6 inhibited high glucose-induced cell apoptosis and renal interstitial fibrosis in mouse renal mesangial cells [31]. Another study demonstrated that low expression of Sox6 declined the activity of Caspase-3 and the percentage of apoptotic cells in mouse P19CL6 cells [34]. In addition, we verified that restored miR-499-5p or depleted Sox6 declined TNF-α, IL-1β and IL-6 contents in BALF and lung tissues of sepsis-induced lung injury mice. Xia etal。 illuminated that TNF-α and IL-6 levels were dramatically raised in sepsis-induced lung injury mice [35]. A study has demonstrated that low expression of Sox6 declines the inflammatory reaction in hippocampal tissues of Alzheimer’s disease [33]. Another result in this study implied that restored miR-499-5p or depleted Sox6 raised SOD, CAT and GSH-Px activities as well as reduced MDA and MPO contents in lung tissues of sepsis-induced lung injury mice. It was presented that CLP-induced ALI was featured by inflammation in morphology, enhanced W/D ratio, raised protein concentration in BALF, higher level of MPO and MDA contents as well as lower level of SOD, GSH-Px and CAT activities in lungs after CLP [36]. It was suggested that the activity of SOD and CAT were decreased in sepsis-induced ALI in mice [37]. Furthermore, a study reported that the overexpressed miR-499-5p and low expressed Sox6 decreased the level of MDA [32].
結論
In conclusion, we found that high expression of miR-499-5p can attenuate the apoptosis of lung tissues cells and inhibit inflammation of sepsis-induced lung injury mice via depleting Sox6, which may have important therapeutic implications in the treatment of sepsis-induced lung injury. Nevertheless, clinical researches are needed to detect the efficacy for the treatment of sepsis-induced lung injury.
Availability of data and material
該当なし。
ナノマテリアル
- 誘電体メタ表面を介した帯域幅の拡大による完全なテラヘルツ分極制御
- 多機能GaN / Feナノ粒子による内皮細胞の標的化
- HepG2腫瘍を有するヌードマウスにおける新規64Cu標識セラノスティックガドリニウムベースのナノプローブの評価
- PEG化リポソームを介したブファリンの改善された抗腫瘍効果および薬物動態
- Ptナノ粒子表面プラズモンとのカップリングによるMgZnO金属-半導体-金属光検出器の大幅な強化
- エッチングストッパーナノレイヤーを介したクリーンなインターフェースプロセスを使用したa-IGZOTFTデバイスのパフォーマンスの向上
- ヒドロキシル基を介したカーボンナノチューブのスライドロール運動モードの調整
- 反応性高周波(RF)熱プラズマによるバイメタルタングステン銅ナノ粒子のその場合成
- 肺および精巣における刷り込み遺伝子の発現および異常なメチル化に対するナノSiO2の影響
- インテグリンαvβ3を標的とするデュアルモードプローブの合成とinvitro研究
- 湿式化学手順によるAuコーティングされたAFMプローブの制御可能な製造