


軟骨形成的に分化しているヒト脂肪由来幹細胞の原子間力顕微鏡ベースのナノスコピー:ナノ構造とインテグリンβ1発現
要約
インテグリンβ1は、分化、移動、増殖、創傷修復、組織発達、および器官形成に関与していることが知られています。インテグリンβ1リガンドと分化クラスター29(CD29)受容体の間の結合確率を分析するために、原子間力顕微鏡(AFM)を使用して、ヒト脂肪由来幹細胞(hADSc)の表面にある天然のインテグリンβ1結合受容体を検出しました。 。インテグリンβ1リガンド-受容体相互作用の結合確率は、2次元細胞培養レベルでの初期の軟骨形成分化中のhADSc上のインテグリンβ1機能化チップによって調べられました。 hADScの細胞形態と超微細構造は、AFMによって測定されました。これは、長い紡錘形の細胞が、軟骨形成誘導中に長さ/幅の比率が減少し、粗さが増加した多角形の細胞になることを示しました。インテグリンβ1リガンドとCD29受容体の結合は、生きているhADScのβ1機能化チップによって検出されました。軟骨形成誘導の0、6、および12日で合計1200の曲線が記録されました。平均破壊力は、それぞれ61.8±22.2 pN、60±20.2 pN、および67.2±22.0pNでした。破裂イベントはそれぞれ19.58±1.74%、28.03±2.05%、33.4±1.89%であり、軟骨形成誘導中にインテグリンβ1リガンドとhADSc表面の受容体の間で結合確率が増加したことを示しています。インテグリンβ1とβ-カテニン/ SOXシグナル伝達経路は、軟骨形成分化中に相関していました。この調査の結果は、AFMが軟骨形成中のhADScへのインテグリンβ1リガンド-CD29受容体結合の変化に対する速度論的および視覚的洞察を提供することを意味します。細胞形態、膜の超微細構造、およびリガンド-膜貫通型受容体結合の確率の変化は、軟骨形成分化プロセスの評価に有用なマーカーであることが実証されました。
背景
変形性関節症(OA)は、高齢者によく見られる変性関節疾患であり[1]、関節軟骨の進行性破壊を特徴とする変性OAを伴います。軟骨は、血管、神経、リンパ組織がなく、高度に組織化されています[2]。細胞外マトリックス(ECM)は主にコラーゲンIIと糖タンパク質で構成されており、軟骨の恒常性にとって非常に重要です。軟骨は無血管であるため、自己複製の能力は限られています。 OA治療(外科的および非外科的の両方)は、OA患者の症状、特に痛みをすばやく緩和することができますが、関節軟骨の正常な構造と機能を回復することはできません[3]。将来的には、治療には、欠陥や変形性関節軟骨を修復するための幹細胞や足場を用いた組織工学が含まれる可能性があります[4]。間葉系幹細胞は、成長因子の組み合わせに応じて、骨形成、脂肪生成、軟骨形成、および筋原性の可能性がある多能性間質細胞です[5]。間葉系幹細胞の分化の分析により、Wnt /β-カテニン、哺乳類のラパマイシン標的(mTOR)、ホスホイノシチド3-キナーゼ(PI3K)、および分化に重要な役割を果たす他の経路が示されています[6,7,8]。ただし、軟骨形成分化が誘導される根本的なメカニズムはとらえどころのないままです。これは、細胞外シグナルが細胞内シグナル伝達経路を活性化するメカニズムに特に当てはまります。我々は、インテグリンβ1が軟骨形成分化中に変化することを発見した。したがって、インテグリンβ1は、さまざまな組織分化シグナル伝達経路に関与しているため、ヒト脂肪由来幹細胞(hADSc)の軟骨形成分化において重要な役割を果たす可能性があると仮定しました。この調査では、Wnt /β-カテニンシグナル伝達経路に焦点が当てられました。
多くの研究は、細胞と細胞外環境との間の相互作用が膜貫通タンパク質、特にインテグリンファミリーメンバーによって調節されていることを示しています[9]。インテグリンは、非共有結合したα鎖とβ鎖のヘテロ二量体膜貫通型糖タンパク質で構成されています[10]。理論的には、64の既知のインテグリンがあり、そのうち24のみが発見されています。インテグリンは、細胞間接着、ECM細胞接着、細胞シグナル伝達、およびアクチン細胞骨格の組織化において重要な役割を果たします[11]。 ECMは組織の恒常性に重要な役割を果たしており、ECMはインテグリンを調節します。インテグリンは、細胞の接着、移動、増殖、分化、細胞死、創傷修復、組織の発達、器官形成など、多くの基本的なプロセスを仲介します。間葉系幹細胞の軟骨形成分化の間、インテグリンβ1の発現はSOXシグナル伝達経路とコラーゲンIIに関連しています。インテグリンβ1ダイマーは軟骨ヘテロダイマーの中で最も顕著なβダイマーであり、多くの異なるαダイマーと相互作用することが知られているため、この調査の焦点はインテグリンβ1ダイマーにありました[12]。分化クラスター29(CD29)は、非常に遅い抗原受容体に関連するインテグリンβ1サブユニットであり、ほぼすべての細胞および組織タイプで発現します。
ここでは、原子間力顕微鏡(AFM)を使用して、hADSc軟骨形成分化中の変化を測定しました。非常に高解像度のタイプの走査型プローブ顕微鏡法として、AFMは、ナノスケールで流体中の単一細胞の形態と細胞膜を検出する新しい機会を提供しました。一方、原子間力顕微鏡(AFM)と組み合わせた単一分子力分光法(SMFS)のシステムを使用して、生細胞上のリガンド-受容体結合を測定しました。 SMFSのシステムは、細胞膜の受容体の変化に対してより敏感であり、結合力の画像が視覚化されました。この研究では、インテグリンβ1リガンド-受容体結合をインテグリンβ1で機能化されたAFMチップによって調べました。 AFMを適用すると、軟骨形成分化がhADSc細胞の形状を変化させ、細胞の粗さを増加させることがわかりました。このアプリケーションは、インテグリンβ1リガンド-受容体相互作用と細胞表面の超微細構造の変化を直接測定することで軟骨形成分化を評価する方法を提供し、細胞表面の調査とスクリーニングを視覚化して改善しました。軟骨形成分化は、膜の組成と構造、および細胞内の細胞骨格の相互作用を変化させます。細胞形態、超微細構造、およびリガンド-膜貫通受容体結合のこれらの変化は、軟骨形成分化メカニズムの評価のための有用なマーカーとして機能します。
メソッド
細胞培養と試薬
この調査では、前述のように3人の外科患者(平均年齢20歳)から細胞を分離しました[13]。すべての患者から情報に基づく同意が得られました。この研究の倫理的承認は、曁南大学第一付属病院から取得されました(補足フォーム)。細胞は、10%熱不活化胎児ウシ血清(FBS、Life Technologies、CA、USA)を添加した低グルコースダルベッコ改変イーグル培地(DMEM、Life Technologies、CA、USA)を含む基本培地で維持されました。 mlペニシリン(Life Technologies、CA、USA)、100μg/ mlストレプトマイシン(Life Technologies、CA、USA)、0.11 mg / mlピルビン酸ナトリウム(Life Technologies、CA、USA)、およびL-グルタミン(Life Technologies、CA、米国)。細胞は、5%CO 2 を含む加湿インキュベーター内で37°Cに維持されました。 培地は3日ごとに交換されます。
インビトロ分化
軟骨形成誘導のために、4〜8継代のhADScを高細胞密度(2×105/10 ml)で播種し、1%FBS、1%インスリン-トランスフェリン-セレン(1%インスリン-トランスフェリン-セレン)を添加したDMEM / F12を含む軟骨形成培地で培養しました。 ITS)+サプリメント(Cyagen、広州、中国)、10 ng / ml形質転換成長因子-beta1(TGF-β1)(Peprotech、ロッキーヒル、ニュージャージー、米国)、100 ng / mlインスリン様成長因子-1( IGF-1)(Peprotech、Peprotech、ロッキーヒル、ニュージャージー、米国)、10-7 Mデキサメタゾン(Sigma、セントルイス、MO、米国)、および50μg/ mlアスコルビン酸(Sigma、セントルイス、MO) 、 米国)。培地は2日ごとに交換され、TGF-β1とIGF-1が新たに追加されました。軟骨形成は、アルシアンブルーおよびトルイジンブルー染色によって評価されました。
骨形成および脂肪生成の分化を誘導するために、4〜8継代の細胞を、それぞれ2週間、骨形成培地および脂肪生成培地で処理しました。骨形成培地は、10-7 Mデキサメタゾン(Sigma、セントルイス、ミズーリ州、米国)、50μg/ mlアスコルビン酸(Sigma、セントルイス、ミズーリ州、米国)、および10 mmol /lβ-グリセロールを添加したDMEMで構成されていました。リン酸塩(Sigma、セントルイス、ミズーリ州、米国)。骨形成はアリザリンレッド染色によって評価されました。
脂肪生成培地は、0.5 mmol / lの3-イソブチル-1-メチルキサンチン(IBMX)(Sigma、セントルイス、ミズーリ州、米国)、1μmol/ lのヒドロコルチゾン(Sigma、セントルイス、ミズーリ州、米国)、 0.1 mmol / lインドメタシン(Sigma、セントルイス、ミズーリ州、米国)。脂肪生成分化はオイルレッドO染色によって評価されました。
フローサイトメトリーによるhADSc表面抗原の同定
hADSCをトリプシンで消化し、DMEMで2回リンスした後、2×10 7 の細胞密度で再懸濁しました。 細胞/ ml。細胞懸濁液(50μl; 1×10 6 細胞)を1.5 mlエポキシエポキシドチューブに加え、抗CD34、抗CD44、抗CD45、抗CD73、抗CD90、抗CD106、抗HLA-DR、および抗CD105抗体とインキュベートしました。暗所で37°Cで20分。抗CD34、抗CD44、および抗CD45は、CST(Beverly、MA、USA)から入手しました。他の抗体はAbcam(Cambridge、MA、USA)から入手しました。次に、細胞懸濁液を×500 g で遠心分離しました。 5分間行った後、上清を除去し、200μlの染色バッファーに細胞を再懸濁します。フローサイトメトリーによる分析の前に、すべてのステップを2回繰り返しました。
イムノブロッティング分析(IB)
以前に記載されたように[14]、免疫ブロッティングのために細胞を収集した。使用した一次抗体は、Abcam(Cambridge、MA、USA)から入手した抗β-カテニン(ab32572)、抗インテグリンβ1(ab30394)、および抗コラーゲンII(ab34712)でした。抗β-アクチン(8H10D10、1:2000)、抗GSK-3β(27C10、1:1000)、および抗SOX(92G2、1:1000)は、Cell Signaling Technology(CST、マサチューセッツ州ビバリー、米国)。二次HRP標識抗体(1:1000–1:3000)はCSTから購入しました。
蛍光抗体法
軟骨形成分化のために、細胞を0、6、および12日間処理し、消化し、24ウェルプレート(Costar353047、コーニング、ニューヨーク、米国)のガラス上で24時間培養しました。細胞を氷冷リン酸緩衝液(PBS)で2回洗浄し、4%パラホルムアルデヒドで室温で15分間固定しました。ブロッキング後、細胞をインテグリンβ1と反応する一次抗体と1時間インキュベートし、続いて暗所でAlexa Fluor 488標識抗マウスIgG(H + L)(CST#4408、マサチューセッツ州、米国)と1時間インキュベートしました。 )、4 '、6-ジアミジノ-2-フェニルインドール(DAPI、Sigma、MO、USA)。ファロイジン染色では、ブロッキング後、細胞を0.2%Triton X-100で30分間透過処理し、次に細胞をDAPIおよびファロイジン-Alexa Flour 573(Life Technologies、CA、USA)で1時間インキュベートしました。 3回洗浄した後、レーザースキャン共焦点顕微鏡(ZEISS、LSM 700、オーバーコッヘン、ドイツ)を使用して、軟骨分化中にインテグリンβ1の細胞内局在と繊維状アクチン(F-アクチン)の変化を評価しました。
AFMのヒントの準備
ばね定数(0.06 N / m)のSi3N4チップ(DNP-10、Bruker Corp)は、次のように抗CD29抗体によって化学的に修飾されました[15]。チップは、アセトン、紫外線、ピラニア溶液(H 2 )で洗浄しました。 SO 4 :H 2 O 2 =3:1、 v / v )さまざまな時間(5分、30分、10分)。精製水で十分にすすいだ後、エタノール中の1%3-APTES(Sigma、St。Louis、MO、USA)の溶液と30分間インキュベートすることにより、チップを形成しました。チップを超純水で3回洗浄し、2.5%グルタルアルデヒド(Sigma、セントルイス、ミズーリ州、米国)溶液で1時間処理しました。過剰なグルタルアルデヒドを水で3回洗浄した。最後に、チップを抗インテグリンβ1溶液(1 mg / ml)に挿入し、4°Cで一晩インキュベートしました。変更されたプローブは、実験前にPBSで洗浄されました。
AFM測定
AFM(Bioscope Catalyst、Bruker、USA)を使用して、軟骨形成分化中のhADScの形態と超微細構造の変化を調査しました。 AFMチップの正確な力の定数はPBSで測定されました。形態と超微細構造を評価するために、細胞をPBSで数回洗浄しました。次に、4%パラホルムアルデヒド溶液を3.5cm 2 に加えた。 15分間の培養皿。細胞をPBSで洗浄した後、使用するまで細胞を4°CでPBSに保存しました。チップのばね定数は、接触モードで4.2〜5.8 N / mの範囲でした。 hADScの形態および超微細構造画像は、AFMによって室温でPBSで撮影されました。 hADScの核を取り巻く超微細構造画像は接触モードで得られました。ナノスコープ分析ソフトウェアを使用して、(0、6、12日目)グループの少なくとも15の異なる細胞の15を超える異なる10×10μm画像の細胞表面の超微細構造を評価しました。インテグリンβ1で修飾されたAFMチップと生きているhADScのCD29受容体との間の結合力を、さまざまな軟骨形成期間(0、6、および12日)で分析しました。結合力は、AFMシステム(Bioscope Catalyst、Bruker、USA)のアプローチ-リトラクトモードで測定されました。インテグリンβ1-生細胞分離イベントを研究するために、インテグリンβ1抗体で修飾されたチップを500 nm / sのアプローチ-リトラクト速度で使用しました。機能化されたチップの力の定数は0.058±0.006N / mでした。セルにかかるしきい値の力は800pNでした。力測定実験の前に、抗インテグリンβ1抗体(100μg/ ml)を30分間細胞に添加しました。インテグリンβ1ブロッキングおよびベアプローブは、インテグリンβ1抗体で修飾されたチップと細胞の間の非特異的破裂力を検出するためのコントロールとしても使用されました。インテグリンβ1リガンド-受容体結合確率の定量化のために、特定の相互作用力曲線をインテグリンβ1抗体で機能化されたプローブによって測定した。 1回の実験で400以上の力曲線が測定され、少なくとも3回の独立した実験の結果が要約されました。したがって、各比較実験で約1200の元の力-距離曲線が、機器のNanoscope分析ソフトウェアを使用して30〜40の異なるセルから取得されました。少なくとも3回の独立した実験の力の値を平均することにより、細胞表面のインテグリンβ1リガンドとCD29受容体の間の相互作用力に対する軟骨形成誘導の効果を決定しました。
逆転写およびリアルタイムPCR
TRIzol®PlusRNAPurificationKits(Life Technologies、CA、USA)を使用し、High Capacity cDNA逆転写キット(Invitrogen)を使用して、製造元のプロトコルにわずかな変更を加えて、1μgのRNAをcDNAに逆転写しました。インテグリンβ1およびGAPDHは、遺伝子特異的プライマーを用いたqRT-PCRを使用して定量化されました。 5'-CTGACTTCAACAGCGACACC-3 '(GAPDH-センス)および5'-CCCTGTTGCTGTAGCCAAAT-3'(GAPDH-アンチセンス)。リアルタイムPCRの場合、ステップ1リアルタイムPCR(Applied Biosystems)は、Fast SYBR @ GREEN Master Mix(Life Technologies、CA、USA)を使用して実行されました。標的遺伝子の発現は、内部標準としてGAPDHに正規化され、比較2-ΔΔCT法を使用して計算されました。各アッセイは3回行った。
統計分析
すべての実験は少なくとも3回行われ、データは平均±標準偏差(SD)として表されました。 2つのグループ間の比較は t によって行われました テスト。グループ平均間の有意差は、一元配置分散分析と、それに続くボンフェローニおよびタムハネのT2検定によって決定されました(等しい分散は想定されていません)。 p の値 <0.05は統計的に有意であると見なされました。
結果と考察
hADScの評価
間葉系幹細胞は、骨形成、脂肪生成、軟骨形成、および筋形成の可能性がある多能性間質細胞です。 hADScを同定するための2つの主要な手段、細胞表面CDマーカーと分化する能力があります[16]。追加ファイル1:図S1および追加ファイル2:図S2に示すように、派生セルはhADScでした。次に、継代3 hADScの細胞増殖をMTTアッセイで測定しました(追加ファイル3:図S3)。
hADSc軟骨形成中に誘発される形態および表面超微細構造の変化
AFMは、細胞の形態と超微細構造をナノスケールで検出するために常に使用されます[17]。細胞の形は、その特殊な細胞機能と組織組織に関係しています。一部の癌研究では、AFMは、薬物効果を評価するための形態学的変化を分析するための高画像技術として使用できます。さらに、間葉系幹細胞の形状は、軟骨形成誘導中に変化します[18]。細胞の形の変化は分化に必要であるように見えますが、細胞の形態が間葉系幹細胞の分化の初期の発達段階に影響を与えるかどうかについてはほとんど知られていません。したがって、hADSc軟骨形成中の形態および膜の超微細構造の変化はAFMによって評価されました。これらの変化は重要であり[19]、細胞の機能に直接影響を与える可能性があるためです[20]。 hADScの表面形態と超微細構造は、さまざまな期間の軟骨形成分化中に調査されました(図1および図2)。形態と表面の超微細構造は、各比較グループで明らかに異なっていました。 0日目、細胞は比較的滑らかな表面を有する細長い紡錘形を有していた。細胞膜の構造は均一でした。軟骨形成誘導後、6日目と12日目に、有意な細胞形態変化が観察されました。ほとんどの細胞は徐々に多角形に収縮し(図1a)、軟骨形成分化中に平均細胞長/幅比が減少しました(図1b)。多くの研究は、細胞形態の変化が細胞の細胞骨格と一致していることを示しています[21]。また、軟骨形成分化中に細胞骨格が変化することもわかりました。これは後者の結果で説明されています。
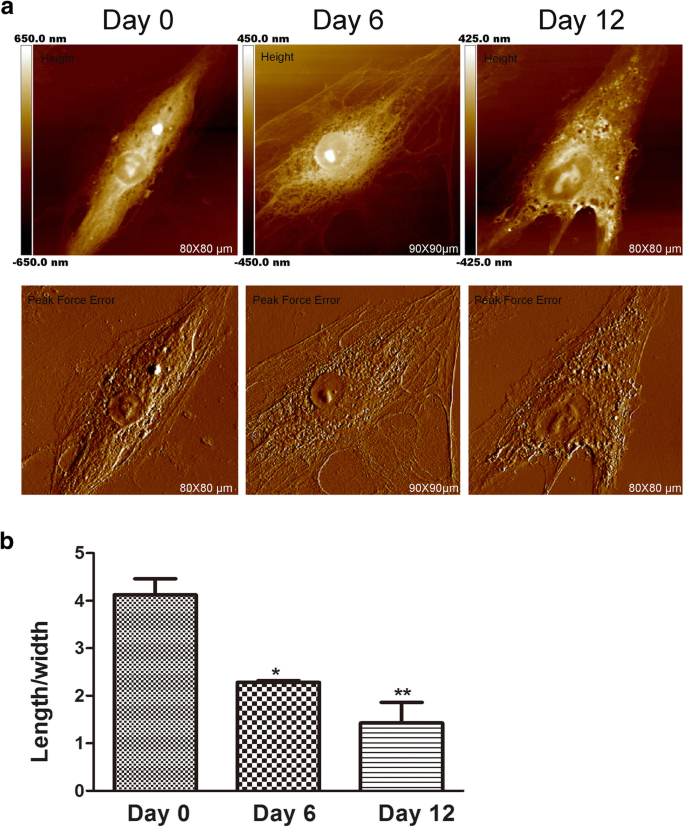
軟骨形成中のhADSc形態の特徴。 a hADSc全体の形態学的画像は、軟骨形成分化の0、6、および12日目に取得されました。画像は、ナノスコープによる高さおよびピーク力誤差画像モデルによって分析された。 b 細胞の平均の長さ/幅の比率は、0、6、および12日の軟骨形成分化処理後に測定されました。 * p <0.05、** p <0.01
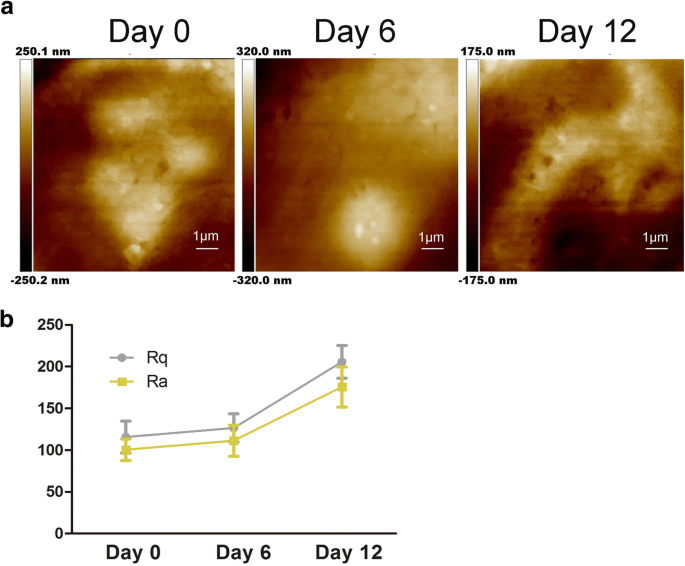
軟骨形成分化中のhADSc膜の超微細構造の特徴。 a 細胞膜の超微細構造の変化は、0、6、および12日間の軟骨形成分化後に評価されました。 b 細胞の表面粗さパラメーターRaおよびRqは、hADScの軟骨形成誘導中に0、6、および12日間測定されました
図2aに示すように、細胞膜の超微細構造も変化しました。粒子が大きくなり、不均一になりました。以前の研究では、RaとRqが、異なる方法で処理された細胞膜の変化を評価するための粗さ値の作成者であることが示されました[22]。 Rqは、二乗平均平方根の粗さについてです。\(\ mathrm {Rq} =\ sqrt {\ frac {\ sum_ {t-1} ^ N {\ left(Zn- \ overline {Z} \ right)} ^ 2 } {N-1}} \); \(\ mathrm {Rq} =\ sqrt {\ frac {\ sum _ {\ mathrm {t} -1} ^ {\ mathrm {N}} {\ left(\ mathrm {Zn}-\ overline {\ mathrm {Z }} \ right)} ^ 2} {\ mathrm {N} -1}}; \)Raは平均粗さ、\(\ mathrm {Ra} =\ frac {1} {N} {\ sum} _ { t-1} ^ N1 \ mid Zi- \ overline {Z} \ mid \)。粗さを取得するために、スキャンサイズは10μm×10μmです。図2bに示すように、2つの異なる領域のRaとRqの両方が、hADScの軟骨形成中に増加しました。 0日目の細胞のRa値とRq値は低く、表面が滑らかであることを示しています(図2b)。 RaとRqの値は、軟骨形成分化と同時に増加し、細胞表面の不均一性が大きく、粗いことを示しています(図2a)。観察された変化に基づいて、軟骨形成分化は細胞形態および細胞の高さ/幅の比率の変化をもたらしました(図1a、b)。 ECMがインテグリンを調節することによって細胞接着を調節できることを示す研究があります[11]。したがって、粗さの値の増加は、軟骨形成中のECMおよび細胞膜の超微細構造の変化を示唆しました。これらのデータは、細胞形態、ECM、および細胞膜構造に影響を与える軟骨形成分化を示しています。
hADScの軟骨形成誘導中の細胞骨格の変化
幹細胞の分化中、細胞の形態と膜の構造変化は、系統特異的な細胞特性の発達に続いて、細胞の細胞骨格に関連しています[21]。図3aに示すように、赤と青の蛍光シグナルはそれぞれF-アクチンとDAPIを示しています。図3aでは、軟骨形成誘導中に細胞骨格が大きく変化しました。一方では、細胞骨格のマイクロフィラメントは0日目のグループで長細胞軸に沿って進みましたが、hADScを軟骨形成分化で12日間処理すると、細胞骨格のマイクロフィラメントは放射状に広がりました。一方、細胞のマイクロフィラメントの分布は、0日目のグループでは均一でしたが、マイクロフィラメントは主に、軟骨形成分化で12日間処理されたhADScの周辺に分布していました。
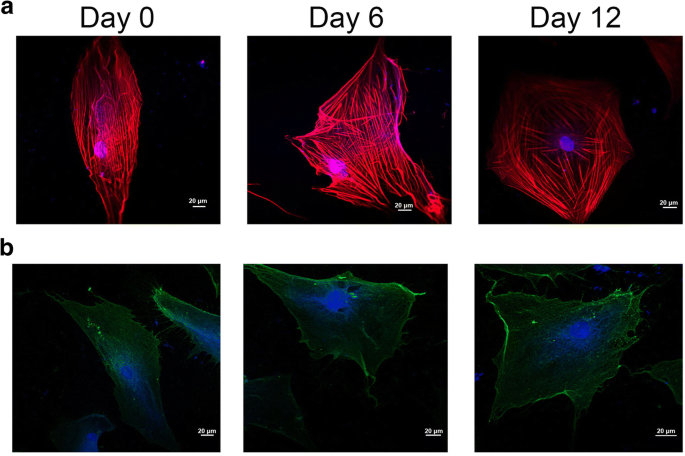
細胞骨格の組織化および軟骨形成的に分化するhADSc上のインテグリンβ1の位置。 a 細胞骨格の変化は、共焦点顕微鏡によるhADScの軟骨形成中に検出されました。 b インテグリンβ1の位置は、共焦点顕微鏡による軟骨形成分化中に測定されました。細胞骨格と核はそれぞれF-アクチンとDAPIで染色されました。赤と青の蛍光シグナルは、それぞれF-アクチンとDAPIを示します
軟骨形成分化により、インテグリンβ1のhADSc上の受容体への結合確率が変化しました
AFMは、リガンドとその受容体の間の結合力を研究するための有用なツールでもあり、細胞表面での膜受容体シグナル伝達を明確にします[23]。 AFMにより、インテグリンβ1とその受容体の間の変化は、視覚的、単純、かつ特定の方法で測定されます。インテグリンβ1リガンド-受容体の生細胞での相互作用は、細胞膜での結合プロセスを探索する方法です。 AFMチップの機能化の手順は、APTESとグルタルアルデヒドの結合によるインテグリンβ1のAFMチップへのカップリングです。これらのヒントは、細胞表面のCD29受容体へのインテグリンβ1の結合を検出するために使用されました(図4a)。単一分子力分光法(SMFS)を使用して、個々の生きているhADScの局所領域内の抗インテグリンβ1生細胞分離力の分布を評価しました(図4b)。代表的な力の曲線を図4c、dに示します。これは、単一分子の曲線(図4c)と2対の破壊ピーク曲線(図4d)を表しています。得られた力曲線の特異性を検証するために、ブロッキング実験と裸のAFMチップ実験を実施しました。ベアAFMチップは、特定の力のピークを検出しませんでした(図4e)。裸のAFM実験は、hADScの表面でのインテグリンβ1リガンド-受容体相互作用の非特異的結合確率が1%未満であることを示しました。ブロッキング実験では、抗インテグリンβ1抗体を細胞と30分間インキュベートした後、インテグリンβ1で機能化したチップを使用して力の曲線を記録しました。ブロッキング抗体は力曲線を90%減少させました(図4f)。抗インテグリンβ1抗体処理後の3つのグループ間で、細胞表面でのインテグリンβ1リガンド-受容体の結合確率に差はありませんでした(図4g)。これらの結果は、抗体で修飾されたAFMチップが力を検出するのに非常に有用であり、インテグリンβ1で機能化されたAFMチップが特異的であることを示しています。
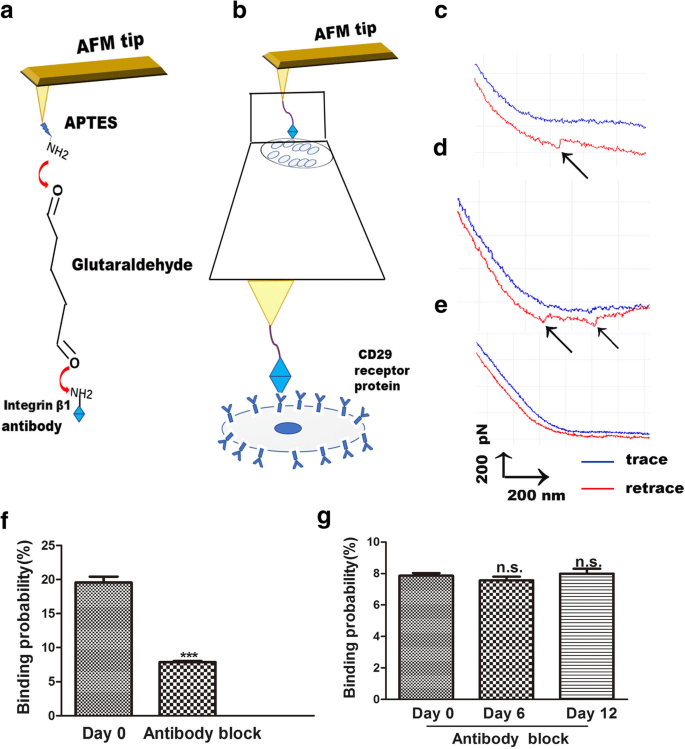
生きているhADSc上のインテグリンβ1官能化AFMチップを用いたAFM力測定。 a インテグリンβ1をAFMチップに固定化するために使用される戦略の概略図。 b インテグリンβ1で機能化されたAFMチップと生きているhADScの間で測定された単一分子力の概略図。 c、d hADScのインテグリンβ1修飾AFMチップで得られた代表的な力曲線、および e システムがインテグリンβ1モノクローナル抗体溶液でブロックされた後。 f 0日目のインテグリンβ1抗体によるブロッキング前後のhADScでのインテグリンβ1機能化チップの結合確率。 g 0、6、および12日目にインテグリンβ1抗体でブロックした後のhADScでのCD29機能化チップの結合確率。 *** p <0.001、n.s ..有意差なし
結合力(破壊力)は、リガンドとその受容体の間の相互作用力です[24]。原形質膜の形態と表面の超微細構造の変化は、分化、アポトーシス、細胞移動など、細胞生物学の多くのプロセスに関連しています。分化中、細胞骨格の変化はインテグリンの変化、特にインテグリンβ1に関連していると考えられています。インテグリンβ1(CD29)は、ECMへの細胞接着および細胞間接着において非常に重要です。また、細胞内タンパク質と相互作用して、アクチン細胞骨格に関連するシグナル伝達分子を刺激することもできます[25]。この研究では、共焦点レーザー走査顕微鏡(CLSM)およびAFMにより、hADSc軟骨形成中に細胞骨格および細胞形態の変化が観察されました。軟骨形成分化の間、細胞骨格、形態、および表面の超微細構造の変化は、細胞状態の新しく信頼できる指標である可能性があります。 CD29受容体であるインテグリンβ1は、蛍光抗体法で判断すると細胞表面に分布しています(図3b)。 hADSc軟骨形成中のインテグリンβ1リガンド-受容体複合体の結合強度と安定性を、分化の0、6、および12日目に評価しました。毎日合計1200の曲線が記録され、平均破壊力はそれぞれ61.8±22.2 pN、60±20.2 pN、および67.2±22.0 pNでした(図5a–c)。力の大きさの分布は、力の平均 + として分析されました。 SD(図5d)。 0日目と6日目で力の平均値に有意差はありませんでした。0日目と12日目で力の平均値に差がありました。結合力の大きさは12日目で増加しました。一方、0、6、12日目の破裂イベントそれぞれ、19.58±1.74%、28.03±2.05%、および33.4±1.89%でした(図5e)。結合確率の増加は、インテグリンβ1(CD29)が軟骨形成分化において重要な役割を果たし、シグナル伝達経路を介して軟骨形成分化の情報を提供する可能性があることも示しています。したがって、軟骨形成分化中のインテグリンβ1ナノドメインの増加は、生きているhADScに対するCD29リガンド-受容体の結合強度に根本的に影響を与える可能性があります。原形質膜の形態および表面超構造の変化は、インテグリンβ1タンパク質構造、コンフォメーション、結合強度、および細胞上のインテグリンβ1リガンド-受容体複合体の安定性の変化を伴いました。要約すると、インテグリンβ1はhADSc軟骨形成分化において必要な役割を果たします。
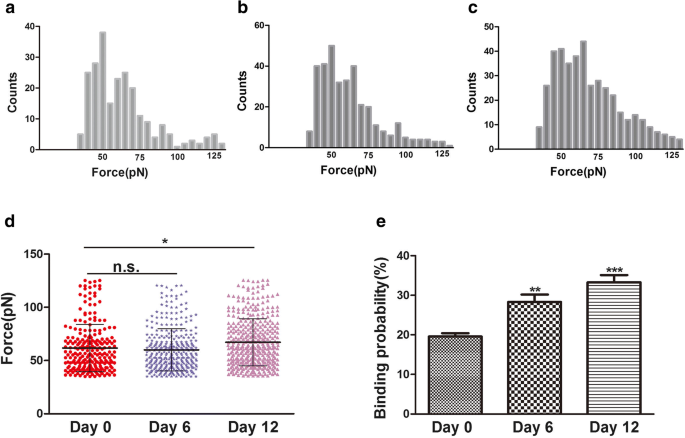
インテグリンβ1で機能化されたAFMチップによって生きているhADScの表面で測定された結合力と結合確率。 a–c 0、6、および12日間のhADSc軟骨形成分化中に得られたインテグリンβ1抗体-受容体の結合力のヒストグラム。 d インテグリンβ1受容体の結合力は、hADScの軟骨形成分化の0、6、および12日目に得られました。 e インテグリンβ1受容体の結合確率は、hADScの軟骨形成分化中に0、6、および12日間検出されました。 * p <0.05、** p <0.01、*** p <0.001、n.s。有意差なし
hADSc軟骨形成分化中のインテグリンβ1のアップレギュレーション
多くの研究は、インテグリンファミリーのメンバーが細胞分化において重要な役割を果たすことを示しています。さらに、インテグリンは細胞外環境と細胞の間の相互作用を調節し、接続されたタンパク質を介したシグナル伝達経路を制御することができます[26]。以前の研究では、結合確率は細胞表面の膜貫通タンパク質(受容体)の密度とコンフォメーションによって影響を受ける可能性があることが示されています[27]。インテグリンコンフォメーションは、リガンドに対する親和性が低い閉じたヘッドピース、またはリガンドに対する親和性が高い開いたヘッドピースの場合があります[28、29]。インテグリンβ1の発現は、軟骨細胞に特徴的なコラーゲンIIの発現が増加することで、転写レベルと翻訳レベルの両方でアップレギュレーションされました(図6a、b)。このように、アップレギュレーションされたインテグリンβ1の発現は、コンフォメーションに関係なく、結合確率の増加と一致していました。
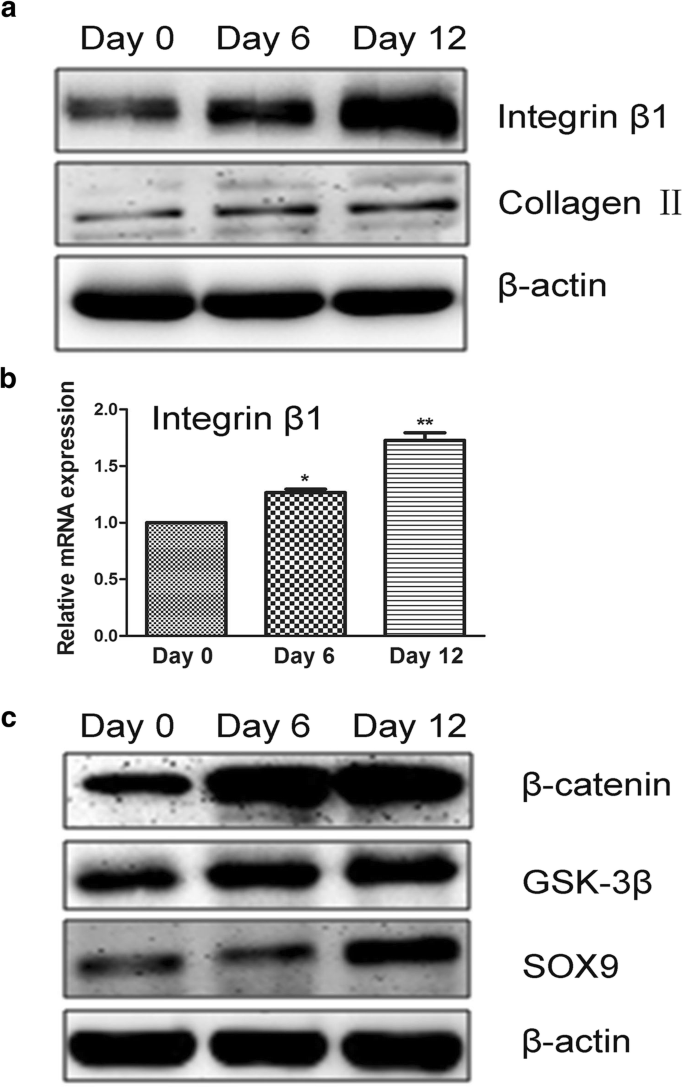
hADSc軟骨形成分化の調節におけるインテグリンβ1およびβ-カテニン/ SOX経路の役割。 a タンパク質インテグリンβ1は、ウエスタンブロッティングによって評価されるように、hADScの軟骨形成中にアップレギュレーションされました。軟骨分化は、異なる日にコラーゲンIIの発現をアップレギュレーションしました。 b インテグリンβ1のmRNAは、hADScの軟骨形成分化中にアップレギュレーションされました。 c 0、6、および12日間のhADScの軟骨形成分化中のβ-カテニン/ SOX経路に関連するタンパク質の測定。 * p <0.05、** p <0.01
以前の研究では、Wnt /β-カテニン、PI3K、およびmTORシグナル伝達経路がインテグリンβ1に関連していることが示されています[30、31、32]。それぞれが間葉系幹細胞の分化に重要です。同様に、研究は、SOXおよびコラーゲンIIがhADScの軟骨形成中にインテグリンβ1によって調節されることを示しました。 SOXは、Wnt /β-カテニンシグナル伝達経路の特徴的な要素です。したがって、我々は、軟骨形成分化がインテグリンβ1を介したβ-カテニン/ SOX経路によって調節されていると仮定した。 SOX、GSK-3β、β-カテニン、およびインテグリンβ1はすべてhADScの軟骨形成中に増加し(図6c)、インテグリンβ1は細胞シグナル伝達を誘導しました。これらのデータは、インテグリンβ1を介してβ-カテニン/ SOX経路によって調節される軟骨形成分化を示しています。
将来と制限
この研究では、細胞形態、膜の構造、およびインテグリンβ1リガンド-受容体の結合確率の変化が、軟骨形成分化プロセスを評価するための有用な画像マーカーであることが実証されました。これは、形態、膜の超微細構造、および軟骨形成分化中の膜貫通タンパク質の変化を評価するための新しい方法です。この研究には限界があります。結合確率の増加はインテグリンβ1の高発現に関連していましたが、軟骨形成中のインテグリンβ1のコンフォメーションは調査されていません。軟骨形成分化中のインテグリンβ1のコンフォメーションを決定するには、さらなる研究が必要です。インテグリンβ1は、hADScの軟骨形成中にβ-カテニン/ SOXシグナル伝達経路に関与することが実証されました。しかし、インテグリンβ1とβ-カテニン/ SOXシグナル伝達経路の関係はまだ完全には確立されていません。この経路におけるインテグリンβ1の正確な役割を特定するには、さらなる研究が必要です。
結論
本研究では、hADScの軟骨形成誘導を評価するために新しい方法(AFM)を採用しました。細胞表面の超微細構造の変化は、AFMイメージングによって評価されました。 AFMを使用して、インテグリンβ1で機能化されたAFMチップにより、インテグリンβ1リガンドとhADScの表面上のその受容体との間の結合力および結合確率を調査しました。 AFMデータに基づいて、軟骨形成中に、細胞の形態は細長い紡錘形から細胞の粗さが増した多角形に変化しました。インテグリンβ1で機能化されたAFMチップを使用することにより、hADScの表面上のインテグリンβ1リガンド-受容体の結合確率と力の大きさが軟骨形成誘導中に増加することがわかりました。イムノブロットにより、インテグリンβ1は、hADScの軟骨形成を調節するβ-カテニン/ SOXシグナル伝達経路に関与することが実証されました。一緒に取られて、これらの結果と確立された方法論は、細胞の形態と粗さのより良い理解に貢献します。さらに、データは、単一分子レベルでのインテグリンβ1リガンド結合プロセスへの熱力学的および速度論的洞察を提供します。このAFM法は、軟骨形成中の生きているhADScのシグナル伝達経路の調査に役立ちます。細胞のナノ構造、膜の構造、および膜貫通タンパク質の結合確率の変化は、軟骨形成分化メカニズムを評価するための有用なマーカーです。このAFM法は、組織工学における間葉系幹細胞の分化のメカニズムを理解するために使用でき、間葉系幹細胞の軟骨形成分化の理解を深めるのに役立ちます。
略語
- AFM:
-
原子間力顕微鏡
- CD:
-
分化のクラスター
- DAPI:
-
4 '、6-ジアミジノ-2-フェニルインドール
- DMEM:
-
ダルベッコの改良イーグル培地
- ECM:
-
細胞外マトリックス
- F-アクチン:
-
フィラメント状アクチン
- FBS:
-
ウシ胎児血清
- hADSc:
-
人間の脂肪を奪われた幹細胞
- IB:
-
イムノブロッティング分析
- IBMX:
-
3-イソブチル-1-メチルキサンチン
- IGF-1:
-
インスリン様成長因子-1
- ITS:
-
インスリントランスフェリンセレン
- mTOR:
-
ラパマイシンの哺乳類標的
- OA:
-
変形性関節症
- PBS:
-
リン酸緩衝液
- PI3K:
-
ホスホイノシチド3-キナーゼ
- SD:
-
標準偏差
- SMFS:
-
単一分子力分光法
- TGF-β1:
-
トランスフォーミング成長因子-beta1
ナノマテリアル
- 卵巣癌細胞の高感度かつ迅速な検出のための柔軟なグラフェンベースのバイオセンサーのデモンストレーション
- マイクロLEDおよびVCSEL用の高度な原子層堆積技術
- 黑リンナノ粒子は、TG2発現のアップレギュレーションを通じてEMSCの骨形成分化を促進します
- スタックカップカーボンナノファイバーの原子および電子構造を明らかにする
- ポリマーナノ構造上のU2OS細胞におけるアクチンと接着斑組織の分析
- ナノテクニックは癌幹細胞を不活性化する
- 有機太陽電池に組み込まれたナノホール型とナノピラー型のパターン化金属電極の比較
- ZnOナノ結晶の合成と逆ポリマー太陽電池への応用
- 原子間力顕微鏡によるポリスチレン薄膜の接着力とガラス転移の研究
- Fe3 +用の再開可能な蛍光プローブBHN-Fe3O4 @ SiO2ハイブリッドナノ構造とそのバイオイメージングへの応用
- 小胞および細胞の周波数変調波誘電泳動:クロスオーバー周波数での周期的Uターン