


内皮細胞相互作用による間葉系幹細胞に対するヒドロキシアパタイトナノ粒子の潜在的な骨誘導効果
要約
ナノヒドロキシアパタイト(ナノHA)は、再生医療の分野で大きな注目を集めています。内皮細胞(EC)-間葉系幹細胞(MSC)の相互作用は骨の再構築に必要ですが、このプロセスでnano-HAが相互作用する方法は不明なままです。ここでは、ECによって媒介される間接的な共培養モデルを使用してMSCに対するHAナノ粒子(HANP)の細胞毒性と骨誘導効果を調査し、根本的なメカニズムを強調しました。細胞毒性以下の用量で、HANPは骨芽細胞遺伝子の生存率と発現、ならびにMSCの石灰化結節とアルカリホスファターゼ産生を増加させることがわかった。これらの現象は、ECによって分泌されるHIF-1αに依存しており、ERK1 / 2シグナル伝達カスケードを引き起こしました。さらに、MSCの骨形成分化に対するHIF-1αの影響を定量的に分析するために、2段階の細胞系統数学モデルが確立されました。これは、HIF-1αが1500 pg / mLまでのMSCの骨形成分化率に対して用量依存的な刺激効果を発揮することを示しており、これは上記の結果と一致していました。私たちのデータは、HANP、EC、およびMSC間の協調的相互作用が骨の再生を刺激するのに役立つ可能性が高いことを示唆しています。さらに、2段階の細胞系統モデルは、骨組織工学におけるエフェクター分子の潜在的な影響を評価するためのinvitroシステムに役立ちます。
はじめに
外傷、先天性奇形、または外科的切除によって引き起こされた骨欠損の再建は、整形外科手術に大きな課題をもたらします[1]。代表的な生物活性セラミックであるヒドロキシアパタイト(HA)が骨代替物として使用されていました[2]。ただし、望ましくない機械的および骨誘導特性は、その臨床応用を制限します[3]。近年、nano-HAは、その独特の生体工学的特性により、より最適なより優れた生物活性と改善された機械的性能を示し、再生医療に関連する生物医学分野で大きな関心を集めています[4]。 nano-HAが骨欠損部に移植されると、骨修復に関与する複数の細胞がそれにさらされます。そのため、nano-HAの生物学的挙動を評価する必要があります。いくつかの証拠は、HAナノ粒子(HANP)がヒト臍帯ウォートンゼリー由来間葉系幹細胞(hWJ-MSC)および骨芽細胞に取り込まれ、骨形成分化が促進されることを直接示しています[5,6,7]。 Dua etal。以前に、人工軟骨の新規軟骨への統合を促進するHANPの能力を報告しました[8]。逆に、HANPはヒト臍帯静脈内皮細胞(HUVEC)の血管新生能を阻害します[9]。人間の健康の観点から、骨の再生に対するHANPの影響をより包括的に理解することが必要であり、人工ナノベースの人工骨の継続的な適用により、そのような研究の緊急性が高まります。
骨の再生は必然的に新生血管の侵入を伴います。 ECは、血管系の内側の細胞内層であり、血液を受動的に送達する役割を果たし、臓器の再生を誘導、特定、誘導し、恒常性と代謝を維持する役割も果たします[10、11]。 MSCは内皮周囲のニッチの一部であり、それらの常駐ニッチ内の特定の生理学的および生化学的微小環境の誘導の下で自己複製および多分化能力を持っています[12]。ツァイ他ECはエンドセリン-1を分泌してMSCを骨および軟骨系統の分化に向けることができることを発見しました[13]。さらに、Saleh etal。マイクロアレイデータ分析を使用して、HUVEC分泌タンパク質と、MSC膜結合受容体と相互作用して増殖と骨形成分化を促進する関連クロストークシグナル伝達経路を特定しました[14]。骨組織工学では、HANPは新生血管と接触し、ECによってエンドサイトーシスを受ける可能性があります。これにより、これらの細胞の生理機能が変化することが示されています[9、15]。これはまた、周囲の骨前駆細胞に影響を及ぼし、パラクリンシグナル伝達を変化させることによって骨の修復に影響を与える可能性があります。ただし、MSCに対するHANPの直接的な影響は調査されていますが、HANPがECを介してMSCの骨形成分化を間接的に誘導できるかどうかについては、まだ明確な理解が不足しています。これは、HANPの影響を理解するために不可欠です。骨の修復に。
この研究では、ECとMSC間の相互作用に対するHANPの生物学的影響についてさらに洞察を得るために、HUVECとhWJ-MSCを使用して間接的な共培養モデルを確立しました。このシステムを利用することにより、HUVECを介したパラクリンシグナル伝達を介したhWJ-MSCに対するHANPの細胞毒性および骨誘導効果を評価しました。 HANPによって誘発される内皮細胞-MSC相互作用に影響を与える重要な要因を特定するために、HANPで刺激されたHUVECの上清中の可溶性因子を、遺伝子レベルとタンパク質レベルの両方で関連するメカニズムに重点を置いて評価しました。結果は、低酸素誘導因子(HIF)-1αがこれらの相互作用において重要な役割を果たすことを示しました。
骨形成のプロセスに対するHIF-1αの影響を定量的に観察および予測するために、2段階の細胞系統とHIF-1αを組み合わせた数学的モデルが確立されました。ここでは、経験的データを分析することにより、2段階の細胞系統モデルを使用して、定義された初期細胞播種密度とHIF-1α濃度に基づいて、任意の時点でのMSC数と分化度を予測しました。初期培養条件とインキュベーション時間について適切な提案を提供します。この研究の結果は、再生医療で使用する革新的な生体材料の開発を促進するのに役立つ、ナノベースの代用骨と生物学的システムとの相互作用に光を当てるのに役立ちます。
材料と方法
粒子の準備と特性評価
20 nm(np20)、20 * 80 nm(np80)のHANP、および純度99.0%以上のマイクロサイズのHA粒子(m-HAP)は、Nanjing Emperor Nano Material Company Ltd(Nanjing、China)から購入しました。粒子のサイズと形状は、透過型電子顕微鏡(TEM; FEI Tecnai G2 Spirit Bio-Twin、FEI、Hillsboro、OR、USA)と走査型電子顕微鏡(SEM; LEO1530VP、Germany)を使用して観察しました。 HA粒子(HAP)の流体力学的サイズとゼータ電位は、Zetasizer NanoZS90とMastersizer3000(Malvern Instruments、Malvern、UK)を介して決定されました。
細胞の準備と培養
すべての実験プロトコルは、南京医科大学の倫理委員会によって承認されました。 HUVECおよびhWJ-MSCは、ドナーからの書面によるインフォームドコンセントを達成した後、前述のように、新鮮なヒト臍帯から採取されました[16、17]。簡単に説明すると、臍帯と臍帯静脈を、1%ペニシリンとストレプトマイシンを含むリン酸緩衝生理食塩水(PBS)(PS; Hyclone、GE Healthcare Life Sciences、Pasching、Austria)ですすいだ。次に、臍帯静脈に0.1%コラゲナーゼI(Sigma、St。Louis、MO、USA)を充填し、37°Cで15分間インキュベートしました。採取後、HUVECはEC培地(ECM)(Sciencell、San Diego、CA、USA)で培養されました。
その後、血管を取り除き、ウォートンゼリーを1 mm 2 にカットしました。 ピースしてから25cm 2 に入れます 組織培養フラスコ(Corning Incorporated、Corning、NY、USA)。これらの細胞は、10%ウシ胎児血清(GIBCO)と1%PSを添加したL-DMEM(GIBCO Life Technology、Grand Island、NY、USA)で培養されました。
hWJ-MSCは、CD13、CD29、CD34、CD44、CD45、CD51、およびCD105(BD Biosciences、San Jose、CA、USA)に対するモノクローナル抗体を使用して表現型を確認するために評価されました。 HUVECは、フォンウィルブランド因子(vWF; Shanghai ChangDao Biotech Co、Ltd.、Shanghai China)を使用して評価されました。これらの実験では、3〜7継代のHUVECと3〜5継代のhWJ-MSCを使用しました。
次に、PBS中の1 mg / mLの粒子懸濁液を、L-DMEMで最終濃度に希釈しました。図1に示すように、HUVECは示された濃度のHAPで18時間インキュベートされました。培地を15,000rpm、4°Cで15分間遠心分離し、10%FBSを添加した上清をhWJ-MSCの馴化培地(CM)として使用して、以下の実験を行いました。 CMは、10 mMのβ-グリセロホスフェート、50μg/ mLのL-アスコルビン酸-2-ホスフェート、および0.1μMのデキサメタゾン(Sigma-Aldrich、米国ミズーリ州セントルイス)を含む骨形成誘導液を添加した骨形成培地で構成されていました。さらに、2-メトキシエストラジオール(2-MeOE2)(Selleck Chemicals、Houston、TX、USA)を特定のHIF-1α阻害剤として使用しました。 2-MeOE2(+)グループでは、hWJ-MSCをHUVECのCMで培養し、HAP補充前に5μMの2-MeOE2で40分間前処理しました。 PD98059を特定のMEK阻害剤として使用しました。 PD98059グループでは、hWJ-MSCは5μMPD98059を含むCMで培養されました。
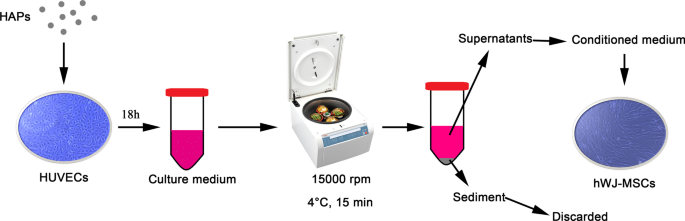
HUVECによって媒介されるHAP / hWJ-MSCの間接共培養の図。略語: HAP ヒドロキシアパタイト粒子、 hWJ-MSCs ヒト臍帯ウォートンゼリー由来の間葉系幹細胞、 HUVEC ヒト臍帯静脈内皮細胞
細胞の生存率と数の決定
細胞生存率は、MTSアッセイキット(MTS; Bestbio、Beyotime Biotechnology、上海、中国)を使用して評価しました。 hWJ-MSCを24時間付着させた後、CMで24時間と72時間培養しました。マイクロプレートリーダー(SpectraMax M2; Molecular Devices LLC、Sunnyvale、CA、USA)を使用して、490nmでホルマザンの吸光度を評価しました。同じ条件下で標準の検量線を使用して、吸光度も細胞数に変換されました。
定量的リアルタイムポリメラーゼ連鎖反応(RT-PCR)
TRIzol試薬(Invitrogen、Carlsbad、CA、USA)を使用して、CMで示された時間インキュベートされたhWJ-MSC細胞から全RNAを単離しました。 T3サーモサイクラー(Mastercycler 5333;エッペンドルフ、ハンブルク、ドイツ)でPrimeScript First Strand cDNA合成キット(TaKaRa、東京、日本)を使用して、1.0 µgのRNAから相補DNAを転写しました。示された遺伝子の発現レベルは、定量的リアルタイム増幅システム(7900HT Fast; Applied Biosystems、Foster City、CA、USA)でFastStart Universal SYBR Green Master(ROX)キット(Roche、Basel、Switzerland)を使用して分析されました。標的遺伝子の相対的なmRNA発現はGAPDHに正規化され、\(2 ^ {{-\ Delta \ Delta C_ {t}}} \)メソッドを使用して決定されました。標的遺伝子のプライマー配列を表1に示します。
<図>アリザリンレッドS(ARS)染色および定量分析
hWJ-MSCを骨形成培地を含む12ウェルプレートで最大14日間培養し、その後、ARS染色(Leagene、Leagene Biotechnology、北京、中国)を使用して細胞外マトリックスの鉱化作用を観察しました。簡単に説明すると、サンプルを無水エチルアルコールで15分間固定した後、1%(w / v)ARS(pH、4.2)で室温で5分間染色しました。染色した細胞を再蒸留水で2回洗浄した後、写真を撮りました。鉱化作用の定量分析のために、300μLの10%(w / v)塩化セチルピリジニウム一水和物(BOMEI、BOMEI Biotechnology、合肥、中国)を各ウェルに添加し、プレートを30分間インキュベートしました。各サンプルからの合計90μLを96ウェルプレートに移し、405nmでの吸光度を3回測定しました。
アルカリホスファターゼ(ALP)染色および定量分析
hWJ-MSCを骨形成培地を含む12ウェルプレートで最大14日間培養した後、BCIP / NBTアルカリホスファターゼ発色開発キット(Beyotime)を使用してALP染色を行いました。簡単に説明すると、hWJ-MSCは4%パラホルムアルデヒドで固定されました。次に、サンプルをニトロブルーテトラゾリウムと5-ブロモ-4-クロロ-3-インドリル-ホスフェートの混合物で4時間染色し、写真を撮りました。 ALP合成を定量化するために、細胞を氷のようなRIPA溶解バッファー(Beyotime)で30分間溶解しました。細胞溶解物を12,000rpm、4°Cで10分間遠心分離し、ALPアッセイキット(Beyotime)を使用して上清をALP定量分析しました。吸光度を405nmで3回測定し、検量線を使用してALP活性に変換しました。
酵素結合免疫吸着測定法(ELISA)
HUVECは2×10 5 で播種されました 細胞/ウェル。 HAPとともに18時間培養したHUVECからCMを収集し(追加ファイル1)、上清をELISA分析にかけました(図1)。ヒトHIF-1αELISAキット(Anhui Joyee Biotechnics、安徽省、中国)を製造元の指示に従って使用しました。
蛍光抗体法
HUVECは12ウェルプレートのスライドに播種されました(7.6×10 4 細胞/ウェル)。 CMに18時間曝露した後、細胞を4%パラホルムアルデヒド(Biosharp、北京、中国)で固定し、PBS中の0.1%Triton X-100(Beyotime)で透過処理した後、1%抗HIF-1α抗体とインキュベートしました[EPR16897 ](ab179483、Abcam、UK)4℃で一晩。続いて、細胞を1%CoraLite594-結合ヤギ抗ウサギIgG(H + L)(Proteintech、USA)と暗所で1時間インキュベートしました。次に、核をDAPI(Beyotime、上海、中国)で染色し、細胞に加えて30秒間反応させました。サンプルは、レーザー共焦点顕微鏡(オリンパス、日本)を使用して検査されました。蛍光強度は、ImageJ v.1.4分析ソフトウェア(ベセスダ、メリーランド州、米国)を使用して定量化されました。
ウエスタンブロット分析
hWJ-MSCをCMで24時間(細胞外シグナル調節キナーゼ(ERK)1/2、p-ERK1 / 2)または骨形成培地で7日間(ラント関連転写因子(RUNX)-2、タイプ1)インキュベートしました。コラーゲン/コラーゲン1(COL I))。次に、細胞をRIPA溶解バッファーで30分間溶解しました。細胞溶解物を遠心分離し、分析のために上清を-20°Cで保存しました。 12%SDS-PAGE後、タンパク質をポリフッ化ビニリデン(PVDF)メンブレンに転写しました。使用した一次抗体は、抗p-ERK1 / 2、抗ERK1 / 2、抗RUNX-2、抗COL I、およびGAPDH(1:1,000、ウサギポリクローナル抗体、Cell Signaling Technology、ボストン、マサチューセッツ、米国)でした。 )。結合していない抗体を除去した後、メンブレンを二次抗体とともに1時間インキュベートしました。膜上の信号は、化学発光ゲルイメージングシステム(LAS4000M; GE Healthcare Biosciences AB、ウプサラ、スウェーデン)を使用して検出されました。 p-ERKとERK、およびRUNX-2 / COL IとGAPDHの比率は、ImageJ v.1.4分析ソフトウェア(Bethesda)を使用して定量化されました。
細胞アポトーシスの評価
hWJ-MSCは10 5 の密度で播種されました 6ウェルプレートのウェルあたりの細胞数。付着細胞を、示された濃度のHIF-1αで示された時間処理した。次に、細胞を収集し、暗所で15分間FITC-アネキシンVおよびPI(Fcmacs、南京、中国)で標識しました。すべてのサンプルは、FACScanフローサイトメーター(BD Bioscience、米国カリフォルニア州サンノゼ)を使用してテストされました。 FlowJo v10(BD Biosciences)を使用してデータを分析しました。
2段階の細胞系統モデル
HAPの大きな可能性と共培養システムの分析の難しさを考慮して、hWJ-の骨形成分化におけるHIF-1αの役割を理解するには、定量分析を提供できる数学的モデルが必要であり、信頼できる予測が必要でした。 MSC。
hWJ-MSCは、0、300、500、1000、1500、2000、3000、4000 pg / mLのHIF-1αと骨形成誘導液で培養されました。これらの濃度でデータをフィッティングした後、コントロール、m-HAP、np80、およびnp20グループのHIF-1α(HUVECによって生成された)の濃度(それぞれ240、300、325、および375 pg / mL)を使用してMATLAB(MathWorks、米国マサチューセッツ州ネイティック)を使用して近似方程式をテストします。モデルを単純化するために、HIF-1αの異なる濃度でのそれらの類似した成長パターンは同一であると考えました。平均微分度を使用して、微分度-時間方程式を適合させた。異なるグループのhWJ-MSCの増殖率、アポトーシス率、および骨形成分化度は、定義された時間に検出されました。
実験データに従って、多段階の細胞系統モデル[18、19]に類似した、簡略化された2段階の細胞系統モデルが確立されました。 C 0 および C 1 それぞれ、hWJ-MSCとターミナルセルのセル番号を表します。 C 0 および C 1 によって管理されています:
$$ \ left \ {\ begin {gathered} \ frac {{{\ text {d}} C_ {0}}} {{{\ text {d}} t}} =\ left [{\ frac {{K --C_ {0} --C_ {1}}} {K} p-(p --1)} \ right] \ upsilon_ {0} C_ {0} \ hfill \\ \ frac {{{\ text {d}} C_ {1}}} {{{\ text {d}} t}} =\ left({2- \ frac {{K --C_ {0} --C_ {1}}} {K} p --p} \ right)\ upsilon_ {0} C_ {0} --AC_ {1} \ hfill \\ \ end {gathered} \ right。$$ここで、 p 、HIF-1αと時間の影響を受け、hWJ-MSCの複製確率を表します。同様に、 d =1 − p は、細胞数の推定データと実験データをフィッティングすることによって得られる分化率です。パラメータv 0 各系統段階(特に v )で細胞がどれだけ速く分裂するかを定量化します =ln2 / c 、ここで、cは細胞周期の持続時間です)。終末細胞のアポトーシス率は A によって象徴されます 。簡単にするために、アポトーシス率は時間の経過とともにわずかに変化することを無視しました。したがって、 A =4.5%は定数です。 K 細胞が無制限に増殖することができなかったため、環境収容力を示します[20]。 HIF-1αはhWJ-MSCの分化率を高め、次のようにモデル化された分化率をもたらします。
$$ \ begin {aligned} d&=\ frac {{d_ {0}}} {{1 +(r * H)^ {m}}} \\ p&=1 --d \\ \ end {aligned} $$ここで、 H HIF-1αの濃度を表します。 d 0 0 pg /mLHIF-1αでの分化率を示します。 rは調節の強度を表します(ここでは、MSCでのHIF-1αの調節の強度を表します)。 mはヒル係数[21]に対応し、MSC分化率とHIF-1α濃度の関係を示します。
統計分析
正規性と等分散性の要件を満たすすべてのデータは、3回以上の独立した実験からの平均±標準偏差(SD)として表されます。 SPSS 24.0ソフトウェア(SPSS Inc.、シカゴ、米国)を使用して、一元配置分散分析または二元配置分散分析を介して統計分析を実行しました。 P 値<0.05は統計的に有意であると見なされました。統計分析は、GraphPad Prism 5(GraphPad Software、カリフォルニア州ラホーヤ)を使用して提示されます。
結果
HAPの特性評価
図2に示すように、HAPは特定のサイズと形状で作成されました。ほぼ球形のnp20の直径は平均20nmで、np80は平均長さ80 nm、幅20nmの棒状でした。 m-HAPもほぼ球形で、直径は約12μmでした。 L-DMEMではすべての粒子が負の表面電荷を持っていました。ゼータ電位の負の値は、骨細胞の付着と増殖、および直接的な骨結合と新しい骨の形成に有意に有利な影響を与えることが示唆されています[22、23]。 L-DMEMで観察された粒子は、水系で凝集する傾向があります。それらの流体力学的サイズもテストされました。これは、それらの生物学的挙動に影響を与える重要な要因でもある可能性があります。

HAPの特性評価。 a のTEM顕微鏡写真 np20および b np80、および c のSEM顕微鏡写真 m-HAP。 d HAPの特性評価( n =6)。略語: TEM 透過型電子顕微鏡、 SEM 走査型電子顕微鏡、 HA ヒドロキシアパタイト、 m-HAP マイクロサイズのHAP粒子
hWJ-MSCに対するHAPの間接毒性
hWJ-MSCに対するHAPの間接毒性を評価するために、MTSアッセイを介して細胞生存率を測定しました。 50 µg / mLのHANPを含むCMは、24時間後と72時間後、特に24時間後にhWJ-MSCの生存率を大幅に刺激する可能性があります。ただし、100 µg / mLのHANPを含むCMは、72時間後のコントロールと比較して細胞生存率を15〜20%低下させました。さらに、25 µg / mLのnp20を含むCMは、24時間後に細胞の生存率を刺激しましたが、np80は刺激しませんでした。これらの現象により、50 µg / mLのHANPが細胞毒性以下の濃度であり、その後のすべての実験で使用されたことが確認されました(図3)。
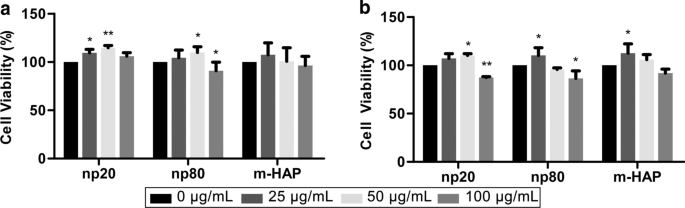
hWJ-MSCに対するHAPの間接毒性。 HAPで間接的に培養されたhWJ-MSCの生存率を a について測定しました。 24および b 72時間。 * P <0.05; ** P <0.01対コントロール。対照群は、HAP処理なしでCMでインキュベートされた細胞で構成され、細胞生存率は対照のパーセンテージとして正規化されました。略語: HAP ヒドロキシアパタイト粒子、 m-HAP マイクロサイズのHAP粒子、 hWJ-MSC ヒト臍帯ウォートンゼリー由来の間葉系幹細胞
hWJ-MSCに対するHAPの間接的な骨誘導効果
hWJ-MSCに対するHANPの間接的な骨誘導効果を特定するために、骨形成遺伝子の発現を定量的RT-PCR分析によって評価しました。 HANPグループのラント関連転写因子2(RUNX-2)、特にnp20の転写レベルは、7日目から21日目まで顕著な増加を示しました(図4a)。 np20グループのI型コラーゲン(Col I)の遺伝子発現は、7日目から21日目まで増強を示し、np80グループは7日目から14日目まで持続的な増加を示しました(図4b)。オステオカルシン(OCN)mRNAは、14日目にHANPグループで明らかにアップレギュレーションされ、骨形成の速度が加速したことを示しています(図4c)。アルカリホスファターゼ(ALP)のmRNAレベルは、HANPグループで明らかに増加し(図4d)、hWJ-MSCの骨形成分化を示しています。ただし、m-HAP群では、対照群と比較して、これら3つの骨形成遺伝子のレベルに関して限られた変化しか見られませんでした(図4)。さらに、多能性マーカーであるNANOG、OCT3 / 4、およびSOX2の発現は、コントロールと比較してHANPsグループで減少しました(図4e–g)。これは、HANPsグループのhWJ-MSCが特にnp20グループ。ウエスタンブロット分析でも同様の結果が得られ(図5c、d)、np20グループがhWJ-MSCでのRUNX-2およびCOLIの発現を間接的に増強できることを示しています。
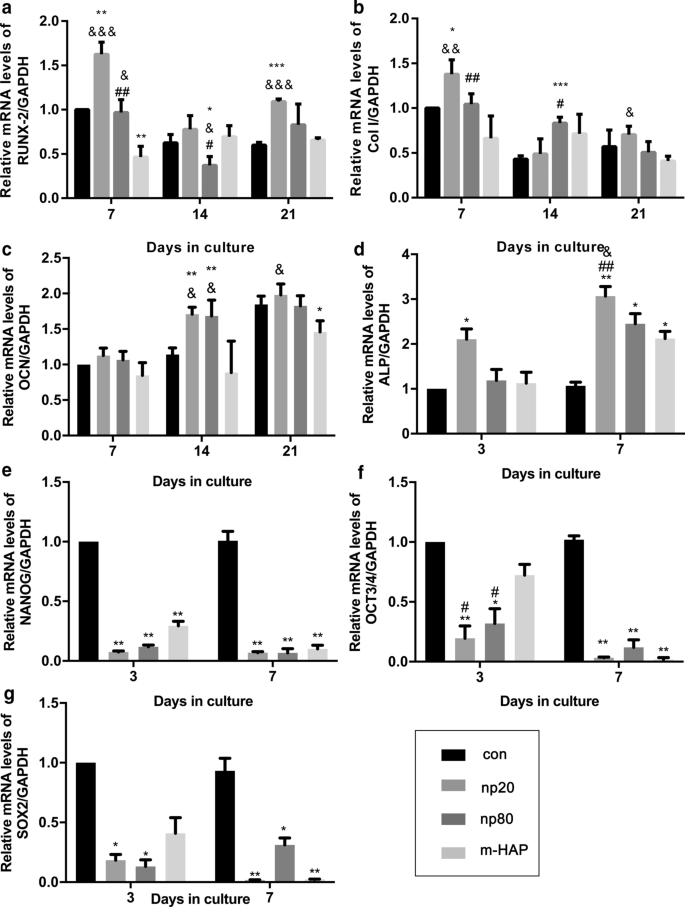
骨形成分化関連遺伝子の発現に対するHAPの間接的効果。 a RUNX-2、 b 列I、 c OCN、 d ALP、 e NANOG、 f OCT3 / 4、および g 示された時間CMで培養されたhWJ-MSCのSOX2遺伝子レベル。 * P <0.05; ** P <0.01; *** P <0.001対対照群; & P <0.05; && P <0.01; &&& P <0.001対m-HAPグループ。 # P <0.05; ## P <0.01対np20グループ。 HAPs処理なしで骨形成培地でインキュベートされた細胞を対照群として使用した。略語: HAP ヒドロキシアパタイト粒子、 m-HAP マイクロサイズのHAP粒子、 hWJ-MSC ヒト臍帯ウォートンゼリー由来の間葉系幹細胞、 RUNX-2 ラント関連の転写因子2、 Col I I型コラーゲン、 OCN オステオカルシン、 ALP アルカリホスファターゼ、 SOX 2 SRY関連のHMGボックス2
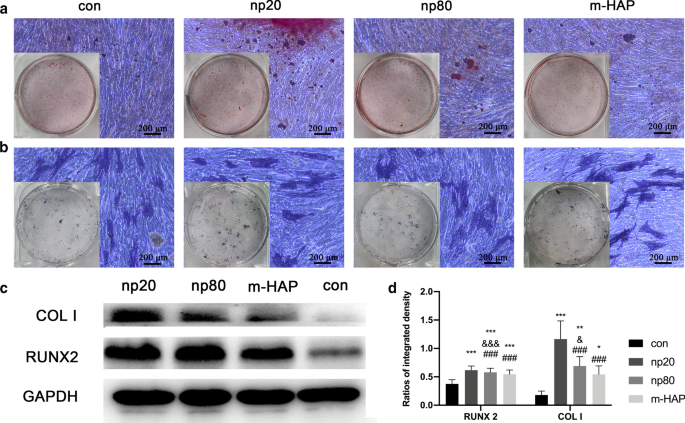
細胞外カルシウム沈着およびALP活性に対するHAPの間接効果。 hWJ-MSCを骨形成培地で14日間インキュベートした。 a 次に、細胞外カルシウム沈着をARS染色によって視覚化しました。 b hWJ-MSCのALP活性は、ALP染色、スケールバー:200μmで評価しました。 c ウエスタンブロット分析は、7日目の骨形成培地におけるhWJ-MSCのRUNX-2およびCOLIの発現を示した。 d パーツ( c )からのRUNX-2およびCOLIのデンシトメトリー測定 )。 HAPs処理なしで骨形成培地でインキュベートされた細胞を対照群として使用した。 * P <0.05; ** P <0.01; *** P <0.001対対照群; & P <0.05; &&& P <0.001対m-HAPグループ。 ### P <0.001対np20グループ。略語: HAP ヒドロキシアパタイト粒子、 m-HAP マイクロサイズのHAP粒子、 hWJ-MSC ヒト臍帯ウォートンゼリー由来の間葉系幹細胞、 ALP アルカリホスファターゼ
hWJ-MSCに対するHANPの間接的な骨誘導効果を視覚的に観察するために、細胞を示された骨形成培地で14日間インキュベートし、続いてARSおよびALP染色を行いました。図5a、bに示すように、m-HAPおよび対照群と比較して、HANP群ではhWJ-MSCの石灰化結節の数の増加とALP活性の上昇が観察されました。さらに、m-HAPは、コントロールと同様に、hWJ-MSCの骨形成分化に対して限定的な効果を示しました。
HAPはHUVECと間接的に共培養されたhWJ-MSCでERK1 / 2シグナル伝達を活性化しました
HAPによるHUVECのパラクリン機能への影響を調査するために、蛍光抗体法およびELISAアッセイを使用して、hWJ-MSCの骨形成分化を促進する可能性のあるタンパク質を特定しました。図6a–cに示すように、HIF-1αの細胞内および細胞外産生はHANP(特にnp20)によって大幅に促進されましたが、m-HAPの産生への影響は限定的でした。
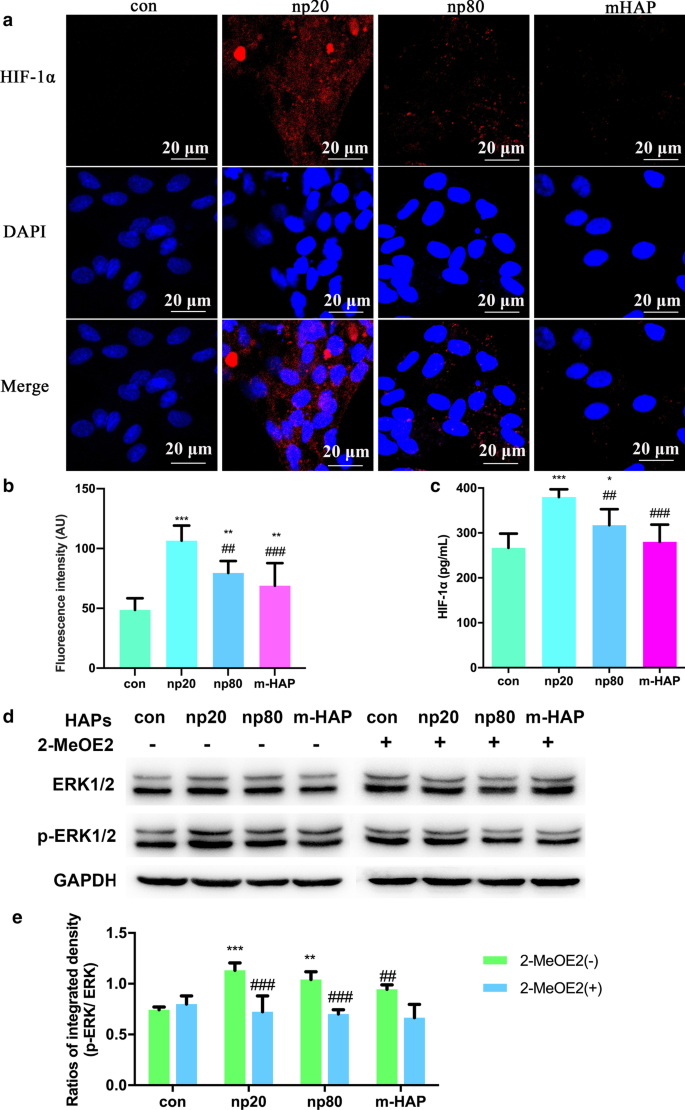
HAPは、HUVECと間接的に共培養されたhWJ-MSCでERK1 / 2シグナル伝達を活性化しました。 a HAPの有無にかかわらず18時間処理されたHUVECで実行されたHIF-1αの免疫蛍光。 b パーツ( a )からのHIF-1αの蛍光強度 )。 c HAPの有無にかかわらず18時間処理されたHUVECの培地中のHIF-1αの細胞外濃度をELISAで測定しました。スケールバー:20μm。 * P <0.05; ** P <0.01; *** P <0.001対コントロール; ## P <0.01; ### P <0.001対np20グループ。 HAP処理なしの細胞を対照群として使用した。 hWJ-MSCはCMで24時間処理されました。 d ERK1 / 2経路における主要なキナーゼの活性化を示すウエスタンブロット分析。 e パーツからのp-ERK1 / 2のデンシトメトリー測定( b )。 ** P <0.01; *** P <0.001対コントロール; ## P <0.01、 ### P <0.001対2-MeOE2(−)グループ。 HAP処理なしでCMでインキュベートされた細胞を対照群として使用した。略語: HAP ヒドロキシアパタイト粒子、 m-HAP マイクロサイズのHAP粒子、 hWJ-MSC ヒト臍帯ウォートンゼリー由来の間葉系幹細胞、 HUVEC ヒト臍帯静脈内皮細胞、 ERK 細胞外シグナル調節キナーゼ、HIF-1α 低酸素誘導因子1α
HIF-1αによって活性化されるhWJ-MSCの分化シグナル伝達経路をより正確に理解するために、ウエスタンブロット分析を介してERK1 / 2経路の主要な調節因子を調べました。図6d、eに示すように、ERK1 / 2全体のタンパク質レベルは変化しませんでしたが、HANPとともに培養したhWJ-MSCではp-ERK1 / 2レベルが明らかに増加し、これは特にnp20グループに当てはまりました。ただし、m-HAPは、HUVECでのHIF-1α産生への影響と同様に、hWJ-MSCでのp-ERK1 / 2レベルにはほとんど影響しませんでした。重要なことに、HIF-1αによって活性化されたhWJ-MSCのp-ERK1 / 2レベルの上昇は、特定のHIF-1α阻害剤である2-MeOE2によってブロックされる可能性があり、HIF-1αがERK1 / 2シグナル伝達経路の上流で機能したことを示していますhWJ-MSCで。
観察されたhWJ-MSC骨形成分化の刺激にHIF-1αが必要かどうかを判断するために、特定のHIF-1α阻害剤(2-MeOE2)をこれらの細胞培養に適用しました。図7に示すように、骨形成培地で培養された2-MeOE2(+)処理グループhWJ-MSCの鉱化マトリックス沈着とALP活性は弱まり、HIF-1αがhWJ-の骨形成分化に不可欠であることを示しています。 MSC。これらの結果に基づいて、HIF-1αによって活性化されるhWJ-MSCの骨形成分化におけるERK1 / 2経路の役割をさらに調査しました。骨形成培地で培養されたhWJ-MSCの鉱化マトリックス沈着とALP活性は、特定のMEK阻害剤であるPD98059の投与後に抑制されました。
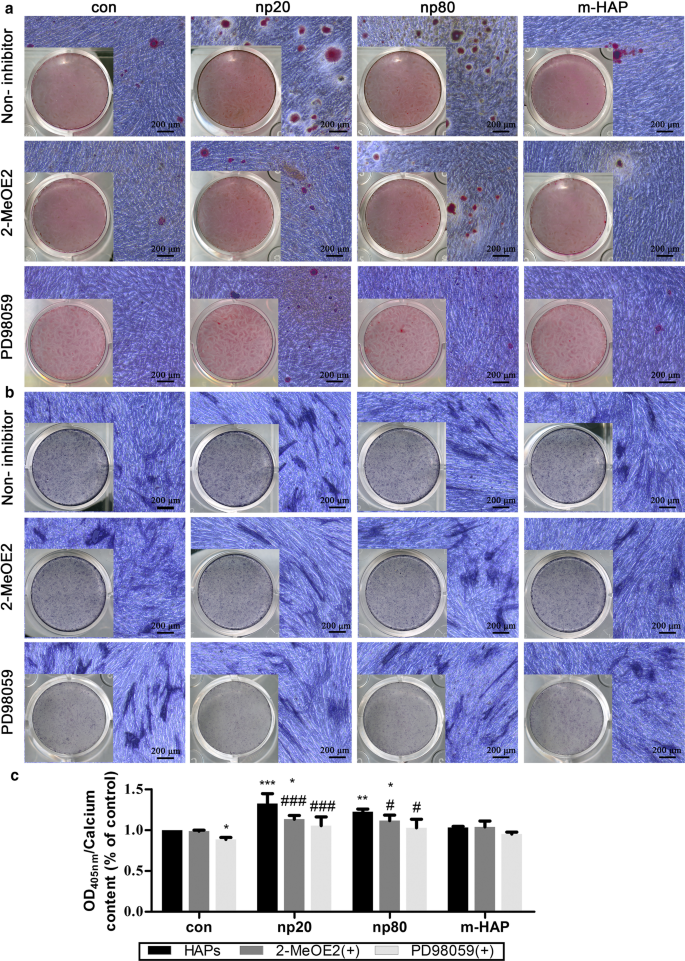
HIF-1αは、ERK1 / 2経路を介してhWJ-MSCの骨形成分化を促進しました。 hWJ-MSCは、PD98059を含むまたは含まない骨形成培地で14日間培養されました。 a 細胞外カルシウム沈着は、ARS染色によって視覚化されました。 b hWJ-MSCのALP活性は、ALP染色によって評価されました。スケールバー:200μm。 c 細胞外カルシウムマトリックスの定量分析。 * P <0.05; ** P <0.01; *** P <0.001対コントロール; # P <0.05; ### P <0.001対np20グループ。 HAPおよびPD98059処理なしの骨形成培地でインキュベートされた細胞を対照群として使用した。略語: HAP ヒドロキシアパタイト粒子、 m-HAP マイクロサイズのHAP粒子、 hWJ-MSC ヒト臍帯ウォートンゼリー由来の間葉系幹細胞、 HUVEC ヒト臍帯静脈内皮細胞、 ERK 細胞外シグナル調節キナーゼ、HIF-1α 低酸素誘導因子1α、 ALP アルカリホスファターゼ
2段階の細胞系統モデル
HIFの濃度とhWJ-MSCの骨形成分化との本質的な関係を定量的に明らかにするために、0〜4000 pg / mLのHIF-1αを使用して8つのグループのhWJ-MSCを処理しました。 2段階の細胞系統数学モデルを使用して、さまざまな濃度のHIF-1αで処理されたこれらのhWJ-MSCの増殖、アポトーシス、および骨形成分化率を分析しました。図8aに示すように、フィッティングデータを使用して、シミュレートされた式を取得しました(\(\ frac {d} {{d_ {0}}} =\ frac {1} {{0.14H ^ {2} -0.43H + 1}} \))および曲線(青い曲線)。HIF濃度の増加に伴い、分化率が最初に増加し、次に減少したことを示しています。具体的には、分化率は1500 pg /mLHIF-1αでピークに達しました。
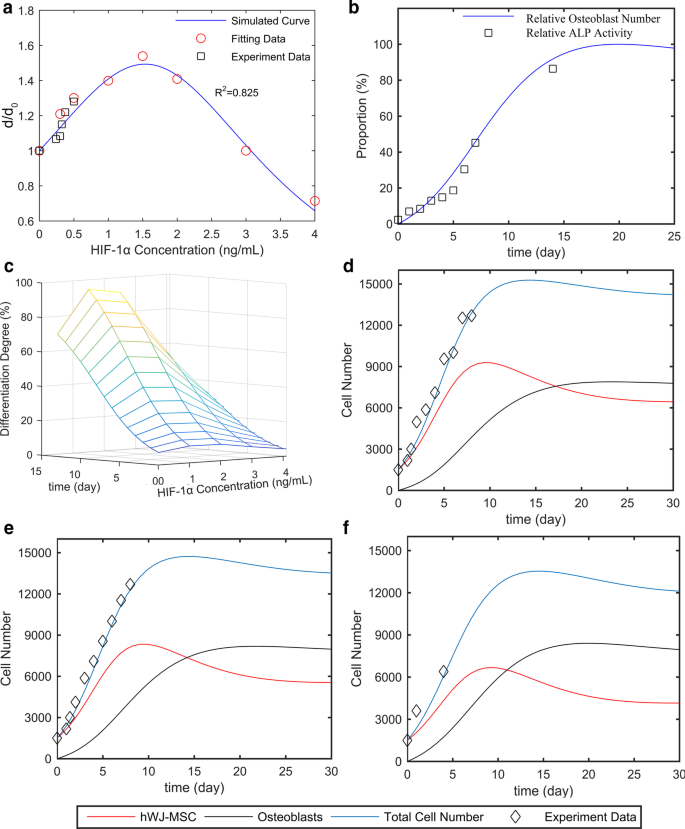
2段階の細胞系統モデル。 hWJ-MSCは、指定された時間、定義された濃度のHIF-1αでインキュベートされました。 a さまざまな濃度のHIF-1α、 b でのhWJ-MSCの相対分化率 異なる培養日の相対ALP活性(hWJ-MSCの分化度)および相対骨芽細胞。 c 時間とHIF-1αとともに進化する分化度の3次元表面。ウェルあたり1500細胞の初期播種密度(96ウェルプレート内)の細胞数、および d 0、 e 375、および f 1500 pg /mLHIF-1α。略語: hWJ-MSCs ヒト臍帯ウォートンゼリー由来の間葉系幹細胞、HIF-1α 低酸素誘導因子1α、 ALP アルカリホスファターゼ
私たちの研究によると、コントロール、m-HAP、np80、およびnp20グループのHUVECによって生成されたHIF-1αの濃度は、それぞれ240、300、325、および375 pg / mLでした。黒い四角は、240、300、325、および375 pg / mLのHIF-1αによって促進される分化率を表しており、これはシミュレートされた曲線とよく一致しています。さらに、異なる濃度のHIF-1αで処理されたhWJ-MSC(相対ALP活性)の分化度は、時間とともに同様に増加しました。したがって、モデルを単純化するために、それらの成長パターンは類似または同一であると見なします。 0日目から7日目までのALP活性の増加は骨芽細胞の数に比例し、骨芽細胞はプラットフォーム期間にピークに達したことがわかりました。したがって、ALP活性を相対骨芽細胞数曲線(骨芽細胞細胞数/最大骨芽細胞数)に適合させた後、ALP活性の最大値を予測し、相対ALP活性(ALP活性/ ALP活性の最大値)を分化度を表す、取得されました(図8b)。これら2つの研究を組み合わせることで、時間とともに進化する分化度とHIF-1αの3次元表面が得られました(図8c)。
最適な培養時間を推定するためには、細胞数をシミュレートする必要がありました。さまざまな濃度のHIF-1αでの分化率を解明した後、各集団のサイズをシミュレートしました。シミュレーションでは、さまざまな濃度のHIF-1αを使用して、ウェルあたり1500細胞(96ウェルプレート内)の初期播種密度を利用しました。図8d–fに示すように、実験データ(黒い四角)は、hWJ-MSCと骨芽細胞の数の合計であるシミュレートされた総細胞数(青い曲線)とよく一致しており、これはこの能力をサポートしています。任意の時点でのhWJ-MSCおよび骨芽細胞の数を予測するためのモデル。さらに、骨芽細胞数は21、18、および15日でプラットフォーム期間に近づき、それぞれ0、375、および1500 pg /mLHIF-1αでした。このモデルは、組織工学を導くための最適な培養時間を提供し、HIF-1αがhWJ-MSCの骨形成分化を促進するという直接的な証拠も提供します。
ディスカッション
ナノバイオマテリアルの最近の進歩により、ナノベースの人工骨代替物は徹底的な調査の領域となっています。蓄積された証拠は、細胞膜に浸透し、内部保持時間を増加させる能力のために、細胞とナノバイオマテリアルの間に複雑な相互作用があることを示唆しています[24、25]。以前の研究では、コラーゲン/アルギン酸ナノフィルムがMSC膜に吸着して、細胞内シグナル伝達カスケードを活性化し、骨形成分化を促進できることが明らかになりました[26]。ウーと彼の同僚によるエレガントな実験は、TiO 2 ナノチューブは、EC-MSC相互作用を介してパラクリン効果と細胞結合を促進することにより、血管新生と骨形成分化を改善することができます[27]。骨組織工学の優れた候補を開発するためには、ナノベースの代用骨と骨修復に関与する細胞との間の直接的なクロストーク、およびそれらの間接的な相互作用を明らかにする必要があります。ただし、これについての現在の理解はまだ限られています。本研究では、間接共培養モデルを利用して、ECによって媒介される間接的な相互作用に関してMSCに対するHANPの生物学的影響をさらに解明しました。
細胞毒性は、ナノバイオマテリアルの生体適合性を評価するための主要な問題です。私たちの以前の研究では、HANPはhWJ-MSCの生存率やアポトーシスに直接影響を与えないことがわかりましたが、他の細胞の媒介を介して異なる影響を与える可能性があります[28]。したがって、HUVECによって媒介されるhWJ-MSCに対するHANPの細胞毒性効果を評価する必要がありました。興味深いことに、CMで24時間および72時間インキュベートした後、hWJ-MSCの生存率は維持され、0〜50 µg / mLのHANPグループ、特にnp20グループでも上昇し、CMにエフェクター分子が存在することを示しています。 HANPの濃度が100µg / mLに達すると、hWJ-MSCに対して細胞毒性を示します。ただし、0〜100 µg / mLのm-HAPは、hWJ-MSCの生存率に影響を与えませんでした(図3)。江ら。特定のサイズの操作されたナノ粒子は、標的細胞機能の調節に関与する下流のシグナル伝達の変化に関連する明確なエンドサイトーシス経路と速度論を持つことができることを示しています[29]。以前の研究では、np20とnp80がHUVECによってエンドサイトーシスされ、これに続いて形態学的変化と大きな液胞が出現し、HUVECの活性化状態を示しました。さらに、np20は、取り込み速度が速く、蓄積が増加しているため、HUVECの活性化が強くなり、パラクリンシグナル伝達を介したhWJ-MSCの生存率が向上する可能性があります。逆に、HUVECによってエンドサイトーシスされるm-HAPはほとんどなく、これがhWJ-MSCの代謝への影響が限られていることを説明している可能性があります[9]。
活性化されたHUVECの潜在的な骨誘導効果をさらに調査するために、50 µg / mLHAPの細胞毒性以下の用量がその後の研究で使用されました。活性化されたHUVECから収集されたCMは、hWJ-MSCでの細胞外カルシウム沈着、ALP活性、骨形成タンパク質の発現、および骨形成遺伝子のmRNA発現を促進しました(図4、5)。骨芽細胞系統の特定に関与する必須の転写因子であるRunx2 [30]は、np20グループで実質的な増強を示し、hWJ-MSCに対する強力な骨誘導効果を示しています(図4aおよび図5c、d)。 np20グループは7日目でCOLI発現の1.5倍の改善を示し(図4b、5c、d)、14日目で2倍の増加を示しました。これは、HANP治療グループに追加の分化骨芽細胞が存在することを意味します(図4b)。 )[30]。 OCNは成熟期の骨マーカーであり[31]、この遺伝子は14日目にHANPグループの有意な増加を示し(図4c)、np20およびnp80がm-HAPと比較して骨の成熟を促進できることを示しています。 ALPは骨芽細胞分化の初期マーカーであり、各グループ、特にnp20グループの培養時間とともに明らかに増加し、MSCから骨芽細胞への追加の形質転換が起こったことを明らかにしています(図4d)。多能性マーカー、NANOG、OCT3 / 4、およびSOX2は、分化の能力を意味します[32]。図4e–gに示すように、HANPグループの遺伝子の発現低下は、HAPグループのhWJ-MSCのほとんどが骨芽細胞に形質転換したことを意味します。
私たちのデータは、HUVECによるHANPのエンドサイトーシスがhWJ-MSCの骨形成分化の改善と関連していることを示しました。ただし、この結果の原因は現在不明です。 HUVECのパラクリン機能の観点から、活性化HUVECの上清中の可溶性分化誘導タンパク質に焦点を当てました。 HIF-1αシグナル伝達は、骨再生中の骨化と血管新生の結合に不可欠です[33、34]。 Heikal etal。負傷したECは正常酸素状態下でもより多くのHIF-1αを分泌すると報告しました[35]。 HANPへの曝露がHUVECの血管新生能力を阻害することも示されています[9]。したがって、CM内のHIF-1αの濃度を測定した結果、HIF-1αの含有量は、m-HAPおよび対照群と比較してHANP治療群で増加したことが示されました(図6a)。 hWJ-MSCの骨形成分化におけるHIF-1αの役割を特定するために、特定のHIF-1α阻害剤である2-MeOE2を使用しました。 HIF-1αの濃度の低下は、これらのhWJ-MSCにおける石灰化マトリックス沈着とALP活性の障害と平行しており、HANPがHUVECのHIF-1α産生を促進してhWJ-MSCの骨形成を促進できることを示しています(図7)。 P>
骨組織工学で使用するためにHANPを適切に適用するには、HANPがHUVECによって媒介されるhWJ-MSCの骨形成分化を促進するメカニズムについてさらに洞察を得る必要があります。 ERK1 / 2経路はHIF-1αの下流にあり[36]、MSCの分化の基本です[37]。この作業では、CMのHIF-1αの濃度はhWJ-MSCのp-ERK1 / 2レベルとよく一致します(図6b、c)。 2-MeOE2を適用した場合、hWJ-MSCのp-ERK1 / 2発現は活性化されず、HIF-1αがERK1 / 2シグナル伝達の上流で機能したことを示しています。 hWJ-MSCの骨形成分化におけるERK1 / 2シグナル伝達の役割に直接取り組むために、特定のMEK阻害剤であるPD98059を使用しました。 ERK1 / 2シグナル伝達の抑制は、hWJ-MSCの最も低い骨形成分化をもたらしました。この発生の考えられる理由の1つは、ERK1 / 2経路が、HIF-1αシグナル伝達とアポトーシスおよび増殖シグナル伝達経路の両方で重要な役割を果たしていることです。 ]。さらに、これは血管内皮増殖因子(VEGF)の存在にも関連している可能性があります。 VEGFはHIF-1αシグナル伝達の下流エフェクターの1つであり[33]、ERK1 / 2経路の活性化を介してMSCの骨形成分化を促進することもできます[37]。私たちの以前の研究では、np20がHUVECでVEGFの産生を誘導することがわかりました[9]。したがって、ERK1 / 2経路の抑制がVEGFの阻害をもたらす可能性があり、これはhWJ-MSCの骨形成分化の低下につながる可能性があります。入手可能な実験結果によると、以下のように要約することができます。 HANPは、m-HAPよりも直接的および間接的な骨誘導効果をより最適に処理することができます。自家骨移植片および骨同種移植片と比較して、HANPの広範な供給源があり、二次的損傷および潜在的な免疫原性はありません。ただし、m-HAPと比較して、HANPはHUVECの血管新生能力を抑制し[9]、時間および用量依存的にわずかな細胞毒性を示します。
最近、増え続ける証拠は、骨の再生におけるHIF-1αの重要性を示しています。ただし、HIF-1α濃度などの特定の初期条件下でMSCの分化率を定量的に予測できる研究はほとんどありません。細胞増殖、アポトーシス、および骨形成分化を考慮に入れて、2段階の細胞系統とHIF-1αを組み合わせた数学的モデルを提示します。これは、実験データと高度に相関しています。 hWJ-MSCの分化率を0〜4000 pg / mLのHIF-1αに適合させることにより、分化率、HIF-1α濃度、および時間を記述するための方程式を取得しました。図8dに示すように、このモデルはさまざまなHIF-1α濃度での細胞数マップを表すことができるため、2段階システムの固有のダイナミクスを調べることができます[40]。さらに、このモデルは、hWJ-MSCの骨形成分化に対するHIF-1αの効果を数学的に検証します。さらに、多段階の細胞系統モデルとロジスティックモデルに基づいて、私たちのモデルは十分に安定しており、人口無制限の爆発の罠に陥ることなく長期予測を可能にします[41]。
既存の実験データを使用することにより、細胞数と分化率の両方を、定義された初期細胞播種密度とHIF-1α濃度で予測することができます。このように、最適なインキュベーション時間も得られます。その結果、HIF-1αの最適濃度を予測し、効率的な組織工学にとって重要な骨形成の最適な時間を決定することができます。 2段階の細胞系統モデルは、2つの細胞系統を持つ幹細胞の増殖と分化を予測するために適用できます。これに基づいて、初期条件と既存の実験データに基づいたモデルを確立して、in vitroでの最適な培養条件を特定できます。これは、invivoでの骨修復の最適化に役立ちます。
結論
この研究では、HUVECによって媒介されるhWJ-MSCに対するHANPの特定の生物学的効果を調査しました。 m-HAPと比較して、np20とnp80はどちらも、時間依存性と用量依存性の両方でわずかな細胞毒性を示しました。重要なことに、HANPのサイズはこの細胞毒性に有意な影響を及ぼさないようでした。私たちのデータは、HANP、特にnp20が、HUVECがHIF-1αのレベルの上昇を分泌するのを促進できることも示しました。これは、ERK1 / 2経路の活性化を介してhWJ-MSCの骨形成分化の増強と直接相関していました(図9)。 。さらに注目すべきことに、2段階の細胞系統モデルの結果は、HIF-1αがhWJ-MSCの骨形成分化率に用量依存的な刺激効果を及ぼすことを示唆しました。さらに、HIF-1αの最適濃度とインキュベーション時間は、in vitroモデルを使用して初期条件に基づいて推定されました。これは、将来、組織工学アプリケーションにとって非常に貴重なものになる可能性があります。まとめると、これらの観察結果は、HANPが細胞間相互作用を調節することによって骨の再生を改善する可能性があるという証拠を提供します。
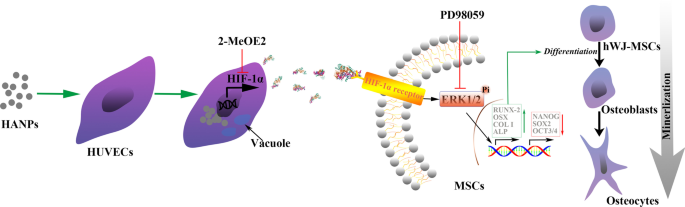
可能なメカニズムの概略図
データと資料の可用性
現在の調査中に使用および分析されたデータセットは、合理的な要求に応じて対応する著者から入手できます。
略語
- HA:
-
ヒドロキシアパタイト
- ハンプ:
-
HAナノ粒子
- hWJ-MSC:
-
ヒト臍帯ウォートンゼリー由来の間葉系幹細胞
- HUVEC:
-
ヒト臍帯静脈内皮細胞
- m-HAP:
-
マイクロサイズのHAP粒子
- PBS:
-
リン酸緩衝生理食塩水
- ECM:
-
ECメディア
- 2-MeOE2:
-
2-メトキシエストラジオール
- ELISA:
-
酵素免疫測定法
- RUNX-2:
-
ラント関連転写因子2
- 列I:
-
I型コラーゲン
- OCN:
-
オステオカルシン
- ALP:
-
アルカリホスファターゼ
- SOX 2:
-
SRY関連のHMGボックス2
- ERK:
-
細胞外シグナル関連キナーゼ
- VEGF:
-
血管内皮増殖因子
ナノマテリアル
- 専用電池
- 太陽電池
- ポリマーナノ構造上のU2OS細胞におけるアクチンと接着斑組織の分析
- ナノテクニックは癌幹細胞を不活性化する
- HT29およびSPEV細胞株に対するAuナノ粒子の影響のinvitro研究
- 磁性ナノ粒子の集合における相互作用効果
- 多機能GaN / Feナノ粒子による内皮細胞の標的化
- 光触媒活性が強化されたAgナノ粒子/ BiV1-xMoxO4の相乗効果
- 6-メルカプトプリンとニューロン透過性ペプチドで修飾された金ナノ粒子によるSH-SY5Y細胞増殖の促進
- 抗菌剤として銀ナノ粒子で装飾された酸化グラフェンベースのナノコンポジット
- インビボでのDNAとのジフェニル-N-(トリクロロアセチル)-アミドホスフェート相互作用に対するC60フラーレンの効果およびinvitroでのヒト白血病細胞株に対するその細胞毒性活性