


鼓室内投与後のラット内耳におけるリポソームナノキャリアの生体適合性
要約
リポソームナノキャリア(LPN)は、低侵襲の鼓室内投与後の高い薬物負荷容量と内耳への効率的な取り込みにより、内耳治療の将来となる可能性があります。ただし、内耳のLPNの生体適合性に関する情報が不足しています。本研究の目的は、鼓室内送達後の内耳におけるLPNの生体適合性を文書化することです。ガドリニウム-テトラ-アザシクロ-ドデカン-テトラ酢酸(Gd-DOTA)の有無にかかわらずLPNは、鼓室内注射によってラットに送達されました。中耳および内耳におけるGd-DOTA含有LPNの分布は、MRIを使用してinvivoで追跡されました。中耳および内耳バリアの機能は、ガドリニウム造影MRIを使用して評価されました。聴性脳幹反応(ABR)を使用して聴覚機能を測定しました。潜在的な炎症反応は、グリコサミノグリカンとヒアルロン酸の分泌、および内耳のCD44とTLR2の発現を分析することによって調査されました。末端トランスフェラーゼ(TdT)を使用してアポトーシスの可能性を分析し、アポトーシス細胞のDNA鎖の遊離3'OH切断をTMR-dUTPで標識しました(TUNEL染色)。その結果、LPNは、鼓室内注射後に効率的に内耳に入りました。 Gd-DOTAを伴うまたは伴わないLPNの鼓室内注射は、中耳および内耳バリアの機能を破壊することも、ラットの聴覚障害を引き起こすこともありませんでした。グリコサミノグリカンとヒアルロン酸の分泌、CD44とTLR2の発現など、内耳の重要な炎症性生物学的マーカーは、LPNの投与による影響を受けませんでした。 LPNの投与に関連する有意な細胞死はありませんでした。 LPNの鼓室内注射は内耳にとって安全であり、LPNは感音難聴の臨床治療におけるドラッグデリバリーマトリックスとして適用される可能性があります。
背景
リポソームナノキャリア(LPN)は、低侵襲の鼓室内投与後の高い薬物負荷容量と内耳への効率的な取り込みにより、内耳治療の将来となる可能性があります[1,2,3,4]。鼓室内アプローチは、メニエール病と突発性難聴の治療のためのクリニックでの以前の戦略であった、非標的領域での治療薬の不必要な蓄積を回避するため、合理的な標的薬物送達アプローチとして耳鼻咽喉科医に広く受け入れられていますゲンタマイシンとコルチコステロイドを使用します。蝸牛におけるモデル治療薬の分子標的化は、特定のペプチド官能化LPNの鼓室内投与によって示されました[5]。さらに、浸透圧ポンプと高性能ポリイミドチューブで構成される新しいデバイスを使用して、中耳から内耳へのLPNの自動持続送達が実現されました[6]。クリニックで最も古いナノ治療プラットフォームとして、LPNは癌、感染症、炎症、痛みなどの治療に安全でした[7,8,9]。ただし、中耳と内耳のLPNの生体適合性は不明なままであり、耳科学に臨床的に適用する前に明確にする必要があります。
耳は外耳、中耳、内耳で構成されており(図1)、鼓室内送達後にLPNにさらされる可能性があります。中耳は最高濃度のLPNにさらされる主要な部位であり、内耳は治療部位であり、危険物質に対して最も敏感な器官であり、外耳道は外耳道から流出する薬剤によって刺激される可能性があります。中耳腔。生物学的障壁は、薬剤の生物学的利用能を制限する最初の防御システムであり、皮膚、粘膜、および神経周囲構造に存在します。内耳のバリアシステムは、内耳の生理的活動に不可欠なイオン恒常性を維持する上で重要な役割を果たします。これらのバリアの機能的変化は、ガドリニウム造影磁気共鳴画像法(Gd-MRI)を使用して正確に評価できます。聴性脳幹反応(ABR)により、聴覚機能の障害を正確に測定できます。したがって、耳(外耳、中耳、内耳を含む)自体がナノ毒性の優れたモデルとして機能します[10、11]。
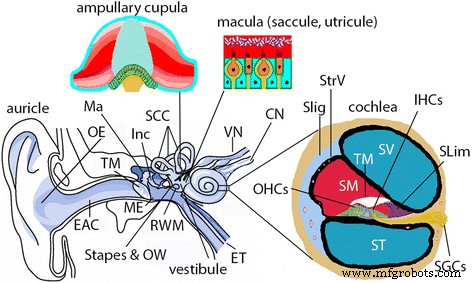
哺乳類の耳のイラスト。哺乳類の耳(人間とラットを含む)は、外耳、中耳、内耳で構成されています。外耳( OE )は、耳介と外耳道( EAC )で構成されています )。中耳( ME )は鼓膜( TM )で構成されています )および槌骨( Ma )を含む耳小骨鎖を収容する空洞 )、砧骨( Inc )、あぶみ骨。中耳腔は、耳管( ET )を介した鼻咽頭の延長です。 )卵円窓( OW )を介して内耳と通信します )および正円窓膜( RWM )。内耳は蝸牛と前庭系で構成されています。蝸牛は聴覚の感覚器官であり、3つのチャンバー、つまり鼓室階( ST )のリンパ管周囲コンパートメントがあります。 )および前庭階( SV )、およびスカラ培地の内リンパ区画( SM )。 SMの側壁には、血管条( StrV )があります。 )およびらせん靭帯( SLig )。 SMの下部には、内有毛細胞( IHC )を含むCortisの器官があります。 )および外有毛細胞( OHC )、蓋膜( TM )、およびスパイラルリンバス(スリム )。らせん神経節細胞( SGC )有毛細胞の機械的電気的伝達に対応する活動電位を発火させ、脳のすべての聴覚入力を供給します。前庭系はバランスに責任があり、3つの半規管( SCC )で構成されています )および前庭。 SCC内のクプラは回転加速度を検出し、前庭の球形嚢と卵形嚢内の黄斑は線形加速度を検出します。 CN 蝸牛神経、 SP スパイラルプロミネンス、 VN 前庭神経、 VS vasspiralis。 (内耳治療におけるZou J. Focal Drug Deliveryから採用:Focal Controlled Drug Delivery。編集者:Domb AJ and Khan W. Springer、London、UK。ISBN:978-1-4614-9433-1、2014; p215- 224)
ヒアルロン酸(ヒアルロン酸)は、天然に存在するポリアニオン性生体高分子であり、基底膜の細胞外マトリックスの主成分です。ヒアルロン酸は、D-グルクロン酸とN-アセチル-D-グルコサミンで構成されており、β-1,4とβ-1,3のグリコシド結合が交互に結合しています。ヒアルロン酸の蓄積は、腎虚血再灌流傷害における透過性の増加と微小循環の炎症に寄与する可能性があります[12]。銀ナノ粒子の耳毒性効果は、以前の報告[11]で、ラット蝸牛におけるヒアルロン酸の蓄積と相関していることが示されました。ヒアルロン酸は組織内のCD44およびトール様受容体2/4(TLR2 / 4)に結合し、生物学的反応を引き起こします[13、14]。ヒアルロン酸に結合する際にCD44によって媒介される生物学的活性は、主に、SRCキナーゼ、Rho GTPase、VAV2、成長因子受容体結合タンパク質2関連結合タンパク質1(GAB1)、アンキリンなどの調節分子およびアダプター分子との相互作用によるものです。およびエズリン[15,16,17]。 CD44はまた、炎症部位へのT細胞の動員と、T細胞を介した内皮損傷の調節に加えて、細胞の取り込みと分解のアプローチを通じてヒアルロン酸の代謝を仲介します[18]。抗内皮細胞抗体による内耳の内皮細胞に対する細胞毒性が、免疫性の突然の感音難聴において血管条損傷を引き起こす一因となる可能性があることが報告された[19]。 TLR2依存性核因子-κBの活性化は、内耳のらせん靭帯線維細胞におけるインフルエンザ菌誘発性単球走化性タンパク質1のアップレギュレーションに関与していると報告されています。これは、慢性中耳炎に続発する内耳機能障害の重要なステップである可能性があります。 20]。 LPNが中耳投与後に内耳障害を誘発する場合、TLR2を介したシグナル伝達経路が重要なメカニズムであるはずです。
鼓室内注射後の内耳におけるLPNの生体適合性を評価することを目的とした。皮膚(外耳道)、粘膜(中耳腔)、および内耳コンパートメントの生物学的バリアの機能は、さまざまな時点でGd-MRIを使用して測定されました。聴覚機能は、ABR測定を使用して評価されました。最後に、グリコサミノグリカンとヒアルロン酸の蓄積、CD44とTLR2の発現、および蝸牛におけるDNA断片化を測定することにより、潜在的な組織病理学的変化を分析しました。
結果
LPNはラット蝸牛の機能変化を引き起こしませんでした
陽性対照群では、蝸牛の外リンパ(Coch)と両側の前庭(Vest)(L、R)の明るい信号(図2a、b)は、Gd-DOTAの取り込みを示しています。銀ナノ粒子(AgNPs)の鼓室内注射後、リンパ管周囲コンパートメントの信号強度は大幅に増加しましたが、外耳道の皮膚、中耳粘膜でも非常に強い信号が検出され、AgNP投与に関連するGd-DOTAの取り込みの強化を示しています(図2a、b)のL(表1)。したがって、評価システムは検証されました。 LPN + Gd-DOTAの鼓室内注射を受けた動物では、注射後3時間で耳小骨鎖、前庭階、鼓室階、前庭の表面に明るい信号が検出され、これらの領域にLPNが明らかに分布していることが示されました(図2c)。 、d)。基底ターンの前庭階の信号強度は鼓室階の信号強度よりも目に見えて強く、現在の動物の卵円窓からのLPNの効率的な侵入を示唆している[21]。注射後6時間で、前庭階と鼓室階の間の信号強度は類似し、蝸牛全体がほぼ均一な信号を示しましたが、前庭にわずかな変化がありました(図2e、f)。ブランクLPNの経鼓膜注射後にGd-DOTAの静脈内注射を受けた動物では、耳小骨鎖の表面にLPNが蓄積していると疑われるブランクLPNの経鼓膜注射を受けた中耳に強い信号があったことを除いて、両側が同様の信号強度を示しました(図。2g、h)。あぶみ骨の中空領域を示す耳小骨チェーンのブラックホール(図2h)。両側の信号強度が等しいことから、LPNの鼓室内注射後、両耳の血液外リンパバリアのGd-DOTAの輸送特性は変化しなかったことが示唆されました(図2g、h)(表1)。
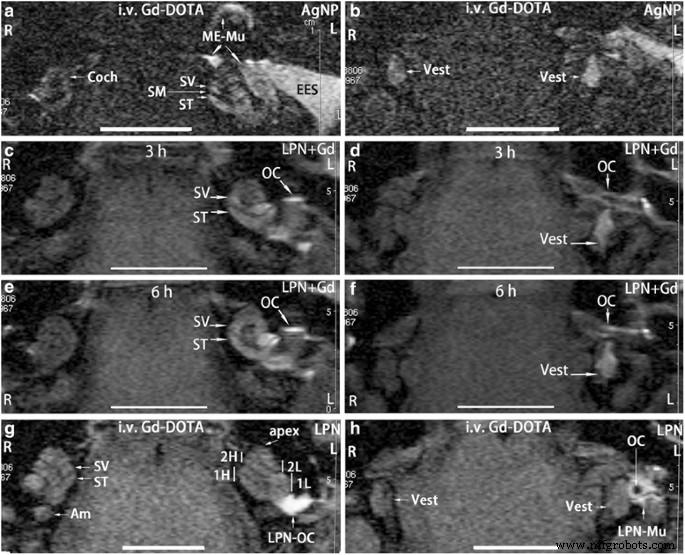
リポソームナノキャリア(LPN)投与後のラット内耳のガドリニウム造影MRI。すべての動物で、ナノマテリアルが左中耳腔の内壁に注入されました。陽性対照は、5時間前に銀ナノ粒子(AgNP)の鼓室内注射に続いてGd-DOTAの静脈内注射の2時間後にラットで画像化されました( a 、 b )。中耳および内耳におけるLPNの動的分布は、 c に示されています。 、 d 、 e 、 f Gd-DOTAの静脈内投与を伴わないGd-DOTA含有LPNの鼓室内注射による。空のLPNが生物学的バリアに与える影響は、5時間前にLPNの鼓室内注射を受けたラットにGd-DOTA(i.v。Gd-DOTA)を静脈内注射してから2時間後にMRIで示されました( g 、 h )。 午前 後部半規管のアンプラ、 Coch 蝸牛、 EES 外耳の皮膚、 L 左耳、 LPN-Mu 中耳粘膜のLPN、 LPN-OC 耳小骨鎖のLPN( OC )、 ME-Mu 、中耳粘膜、 R 右耳、 SMスカラメディア 、 ST 鼓室階、 SV 前庭階、ベスト 前庭、 1H 蝸牛の基底高回転、 1L 蝸牛の基底下部ターン、 2H 蝸牛の2番目に高いターン、 2L 蝸牛の2番目の低いターン。 スケールバー =5mm
LPN + Gd-DOTAもLPNも、重大な難聴を引き起こしませんでした。これは、2、4、8、16、および32 kHz、2、4、および32kHzの周波数でクリックおよびトーンバーストの刺激を使用して測定されたABRしきい値シフトとして表されます。投与後7日、脱イオン水の鼓室内注射を受けた耳と比較して(dH 2 O)(図3)。
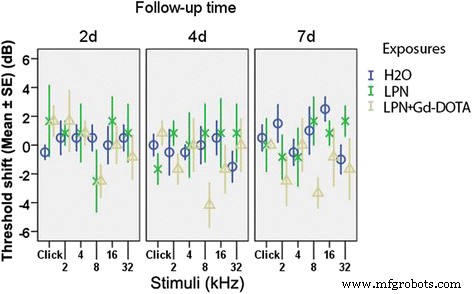
聴性脳幹反応によって測定されたラットの聴覚機能に対するリポソームナノキャリアの鼓室内注射の影響。難聴は閾値シフトとして表されました。グループ間でわずかな違いがありました( p > 0.05、一元配置分散分析)。 n =各グループで6。 H2O 陰性対照群 LPN における脱イオン水の鼓室内注射 空のリポソームナノキャリア、 LPN + Gd-DOTA Gd-DOTAを含むLPN、 2d 、 4d 、および7d 注射後2、4、7日
LPNはラット蝸牛にグリコサミノグリカンの蓄積を誘発しませんでした
ヘマトキシリンおよびエオシン染色では、LPNが通過するアブミ骨および卵円窓を含むすべての分析動物の蝸牛における白血球およびフィブリンの炎症性浸潤は示されませんでした(図4)。過ヨウ素酸シッフ染色は、dH 2 > O.基底ターンから頂点までの信号強度に勾配のある増加があり、その差は血管条で有意でした(図5および6)。 LPNおよびLPN + Gd-DOTAの鼓室内注射を受けた動物では、蝸牛の信号勾配は変化しませんでした(図5および6)。
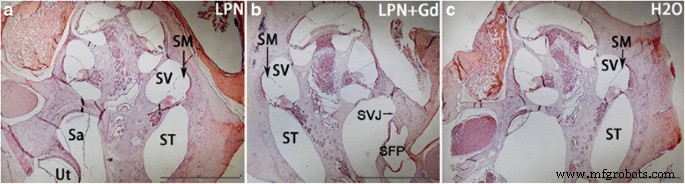
リポソームナノキャリアに曝露されたラット蝸牛のヘマトキシリン-エオジン染色。 LPNの投与を受けた蝸牛には炎症性浸潤はありませんでした( a )、LPN + Gd( b )、およびH2O( c )。 円で囲まれた領域 強度測定のための関心領域の選択を示しました( a )。 LPN 空のリポソームナノキャリア、 LPN + Gd Gd-DOTAを含むLPN。 Sa 球形嚢、 SFP あぶみ骨のフットプレート、 SVJ stapediovestibular関節、 SM スカラメディア、 ST 鼓室階、 SV 前庭階、 Ut 卵形嚢。 スケールバー =1 mm
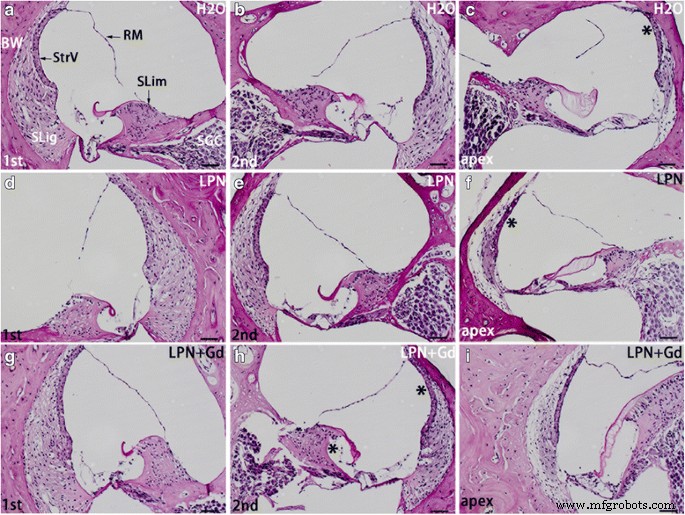
リポソームナノキャリアに曝露されたラット蝸牛におけるグリコサミノグリカン分泌は、過ヨウ素酸シッフ染色光学顕微鏡を使用して検出されました。スパイラルリンバス( SLim )と骨の壁( BW )の蝸牛は、陰性対照( H2O )のグループで最も強い染色を示しました。 )( a – c )、空のリポソームナノキャリア(LPN)( d – f )、およびGd-DOTAを含むLPN( LPN + Gd )( g – i )。明らかに高い強度の染色領域は、 c の*で示されました。 、 f 、および h 左の列と比較して。 RM ライスナーの膜、 SGC 蝸牛神経節細胞、 SLig らせん靭帯、 StrV 血管条、 1st 基礎ターン、 2番目 2ターン目。 スケールバー =50μm
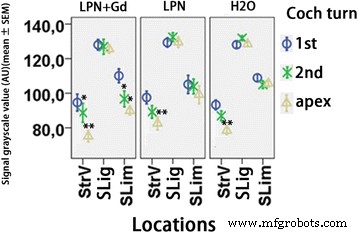
過ヨウ素酸シッフ染色を使用して検出されたリポソームナノキャリアに曝露されたラット蝸牛におけるグリコサミノグリカン分泌の定量化。 n =各グループで3。 AU 任意単位、 H2O ネガティブコントロール、 LPN 空のリポソームナノキャリア、 LPN + Gd Gd-DOTAを含むLPN、 SLig らせん靭帯、スリム スパイラルリンバス、 StrV 血管条、 1st 基礎ターン、 2番目 2ターン目。 * p <0.05、** p <0.01(事後分析として使用されるLSDテストを使用した一元配置分散分析)
LPNによるラット蝸牛のヒアルロン酸分泌に軽微な影響がありました
dH2Oの鼓室内注射を受けたラットの蝸牛では、ヒアルロン酸の陽性染色が主に蝸牛神経節細胞、外ラセン溝細胞、毛細血管内皮細胞などで検出されました(図7)。基底および第2ターンのらせん靭帯線維細胞の信号強度は、頂点の信号強度よりも有意に高かった。これらの違いは、LPNとLPN + Gd-DOTAを適用したラットの蝸牛では重要ではなくなり、らせん靭帯線維細胞によるヒアルロン酸の分泌がLPN投与の影響を受けたことを示しています(図8)。 LPN + Gd-DOTAはまた、基底ターンのらせん靭帯線維細胞の染色を減少させました。ただし、LPNおよびLPN + Gd-DOTAの鼓室内注射による蝸牛細胞の大部分でのヒアルロン酸の分泌への影響はありませんでした(図7および8)。
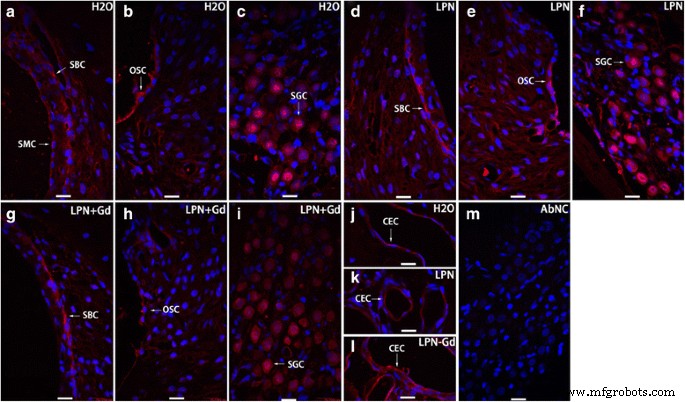
リポソームナノキャリアに曝露されたラット蝸牛におけるヒアルロン酸分泌は、免疫蛍光共焦点顕微鏡で検出された。線条基底細胞( SBC )に陽性染色が見られた )、外ラセン溝( OSC )、らせん神経節細胞( SGC )、および毛細血管内皮細胞( CEC )ネガティブコントロール( H2O )のグループのモダイオラス )( a – c 、 j )、空のリポソームナノキャリア( LPN )( d – f 、 k )、およびGd-DOTAを含むLPN( LPN + Gd )( g – i 、 l )。ネガティブコントロール( AbNC )を省略した抗体には染色はありませんでした )( m )。 CEC 毛細血管内皮細胞、 ISC 内ラセン溝、 SBC 線条体基底細胞、 SL-I らせん靭帯線維細胞I型、 SLSF スパイラルリンバス衛星線維細胞、 SMC 血管条辺縁帯細胞。 スケールバー =16μm
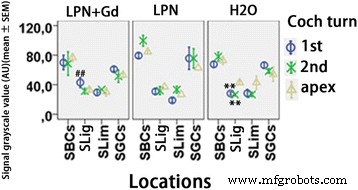
免疫蛍光共焦点顕微鏡を使用して検出されたリポソームナノキャリアに曝露されたラット蝸牛におけるヒアルロン酸分泌の定量化。 n =各グループで3。 AU 任意単位、 H2O ネガティブコントロール、 LPN 空のリポソームナノキャリア、 LPN + Gd Gd-DOTAを含むLPN、 SBC 線条体基底細胞、 SGC らせん神経節細胞、 SLig 、らせん靭帯、スリム スパイラルリンバス、 1st 基礎ターン、 2番目 2ターン目。 ** p <0.01(頂点と比較して)、## p <0.01(基底ターンのLPNおよびH2Oグループと比較)(事後分析として使用されるLSDテストを使用した一元配置分散分析)
LPNはラット蝸牛のCD44細胞集団を変化させませんでした
dH 2 にさらされた蝸牛 O、線条体中間細胞、線条体基底細胞、らせん靭帯線維細胞、らせん神経節細胞、コルチ器のDeiters細胞、およびモディオラスとらせん靭帯の毛細血管内皮細胞は、CD44の強い染色を示しました。蝸牛ターン間で信号強度にわずかな違いがありました。 CD44陽性集団と発現強度は、LPN + Gd-DOTAまたはLPNのいずれの鼓室内注射の影響も受けませんでした(図9および10)。
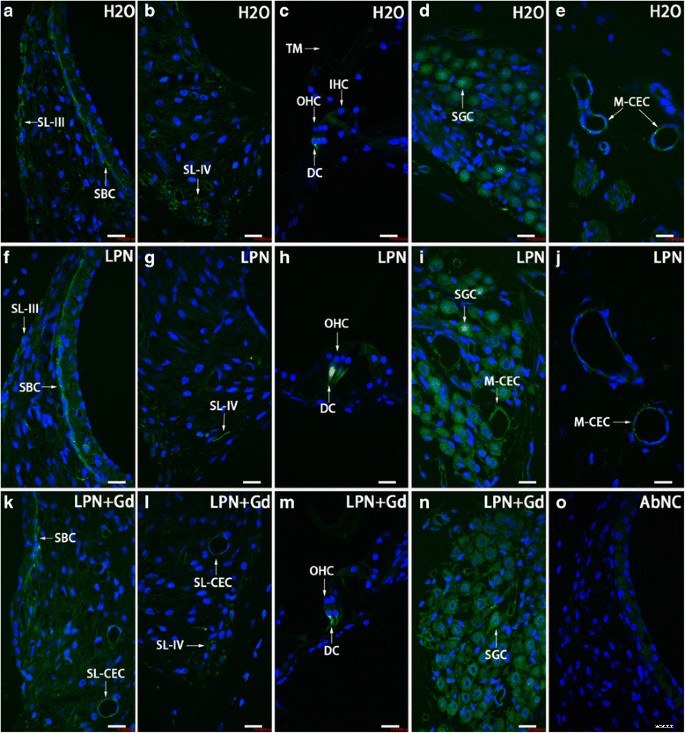
リポソームナノキャリアに曝露されたラット蝸牛におけるCD44陽性細胞分布は、免疫蛍光共焦点顕微鏡を使用して実証されました。 CD44陽性細胞は主に線条体基底細胞( SBC )で検出されました )、らせん靭帯( SL )、ディーターの細胞( DC )、らせん神経節細胞( SGC )、および毛細血管内皮細胞( CEC )ネガティブコントロールのグループ( H2O )( a – e )、空のリポソームナノキャリア( LPN )( f – j )、およびGd-DOTAを含むLPN( LPN + Gd )( k – n )。ネガティブコントロール( AbNC )を省略した抗体には染色はありませんでした )( o )。 IHC 内有毛細胞、 M-CEC モディオラスの毛細血管内皮細胞、 SL-CEC らせん靭帯の毛細血管内皮細胞:らせん神経節細胞、 SL-III らせん靭帯線維細胞タイプIII、 SL-IV らせん靭帯線維細胞タイプIV、 TM 蓋膜、 OHC 外有毛細胞。 スケールバー =16μm
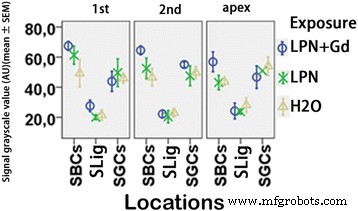
免疫蛍光共焦点顕微鏡を使用して検出されたリポソームナノキャリアに曝露されたラット蝸牛におけるCD44タンパク質レベルの定量化。グループ間でわずかな違いがありました( p > 0.05、一元配置分散分析)。 n =各グループで3。 n =各グループで3。 AU 任意単位、 H2O ネガティブコントロール、 LPN 空のリポソームナノキャリア、 LPN + Gd Gd-DOTAを含むLPN、 SBC 線条体基底細胞、 SGC らせん神経節細胞、 SLig らせん靭帯、 1st 基礎ターン、 2番目 2ターン目
LPNはラット蝸牛のTLR2発現を変化させませんでした
dH2Oに曝露された蝸牛では、線条体基底細胞、らせん靭帯線維細胞、根細胞、らせん神経節細胞、コルチ器官の柱状細胞、およびモディオラスの毛細血管内皮細胞がTLR2に対して強い染色を示した。蝸牛ターン間で信号強度にわずかな違いがありました。 TLR2陽性集団と発現強度は、LPN + Gd-DOTAまたはLPNのいずれの鼓室内注射の影響も受けませんでした(図11および12)。
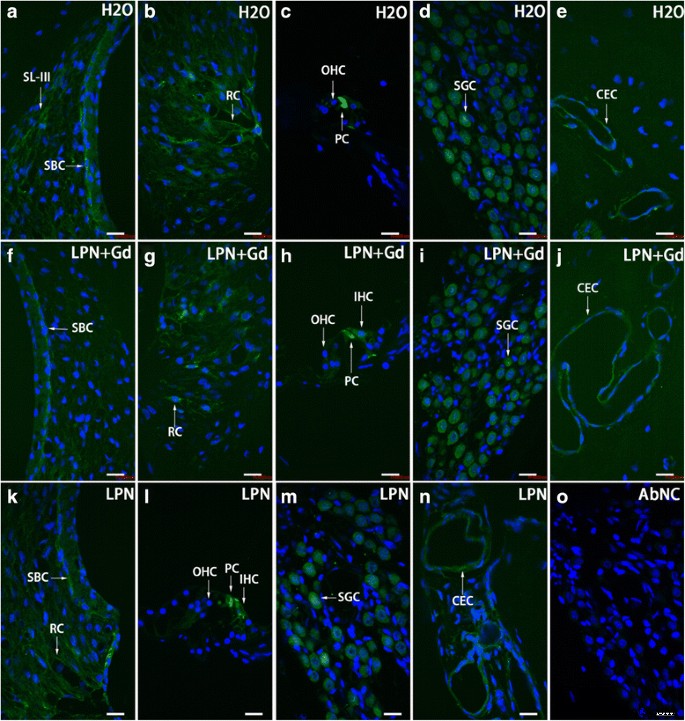
リポソームナノキャリアに曝露されたラット蝸牛におけるTLR2陽性細胞分布は、免疫蛍光共焦点顕微鏡を使用して実証されました。 TLR2陽性細胞は、主に線条体基底細胞( SBC )で検出されました。 )、らせん靭帯( SL )、ルートセル( RC )、ピラーセル( PC )、らせん神経節細胞( SGC )、および毛細血管内皮細胞( CEC )ネガティブコントロールのグループ( H2O )( a – e )、空のリポソームナノキャリア( LPN )( k – n )、およびGd-DOTAを含むLPN( LPN + Gd )( f – j )。抗体を省略したネガティブコントロール( AbNC )には染色はありませんでした。 )( o )。 IHC 内有毛細胞、 SL-III らせん靭帯線維細胞III型、 OHC 外有毛細胞。 スケールバー =16μm
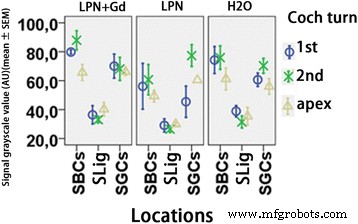
免疫蛍光共焦点顕微鏡を使用して検出されたリポソームナノキャリアに曝露されたラット蝸牛におけるTLR2タンパク質レベルの定量化。グループ間でわずかな違いがありました( p > 0.05、一元配置分散分析)。 n =各グループで3。 AU 任意単位、 H2O ネガティブコントロール、 LPN 空のリポソームナノキャリア、 LPN + Gd Gd-DOTAを含むLPN、 SBC 線条体基底細胞、 SGC らせん神経節細胞、 SLig らせん靭帯、 1st 基礎ターン、 2番目 2ターン目
LPNはラット蝸牛で細胞死を引き起こしませんでした
未処理のラットの蝸牛にランダムに分布するまばらなアポトーシス細胞がありました。驚いたことに、あぶみ骨と卵円窓のニッチのフットプレートにはアポトーシス細胞が豊富にありました。 LPNおよびLPN + Gd-DOTAの投与によるアポトーシス細胞の量および分布パターンへの影響はありませんでした(図13)。
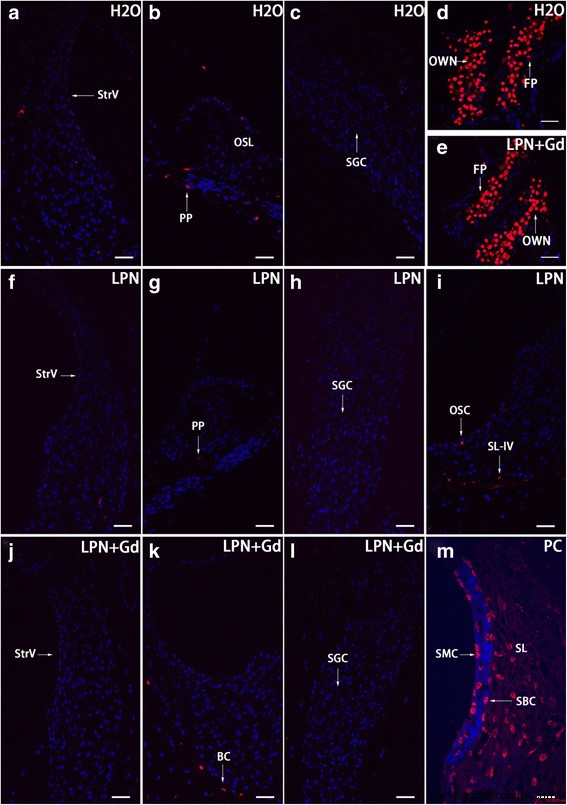
リポソームナノキャリアに曝露されたラット蝸牛のアポトーシスは、TUNEL染色共焦点顕微鏡を使用して実証されました。アポトーシス細胞は、ネガティブコントロール( H2O )のグループのラットの蝸牛細胞でまばらに検出されました。 )( a – c )、空のリポソームナノキャリア( LPN )( f – i )、およびGd-DOTAを含むLPN( LPN + Gd )( j – l )。あぶみ骨のフットプレートにはアポトーシス細胞が豊富にありました( FP )および卵円窓のニッチ( OWN )両方のグループ( d 、 e )。ポジティブコントロール(PC)では、らせん靭帯線維細胞( SL )で豊富なTUNEL染色が検出されました。 )、線条体基底細胞( SBC )、および線条体辺縁細胞( SMC )。 PP 蝸牛神経節細胞の周辺突起( SGC )、 OC 骨細胞、 OSC 外ラセン溝、 SL-IV らせん靭帯線維細胞のIV型、 SLim スパイラルリンバス、 StrV 血管条。 スケールバー a – l =32μm、 m =16μm
ディスカッション
LPNは、LPN内にカプセル化された薬物模倣薬としてGd-DOTAを使用したMRIによって実証された、鼓室内注射後に効率的に内耳に入りました(図2c–f)。以前の研究では、正円窓が内耳に入るLPNの主要な経路であることが示されていましたが[6]、今回の観察では、正円窓よりも正円窓の経路が中耳から内耳にLPNを輸送するのに効率的であることが示されました。耳。この結果は、両方の経路が、標的とされた中耳内壁投与後のLPNの内耳負荷において重要であることを示唆しました。ガドリニウム増強内耳MRIの最も効率的なinvivo法を使用して、生物学的バリアを評価し、周波数特異的ABRを使用して聴覚機能を評価し、本研究は、LPNおよびLPN + Gd-DOTAの鼓室内注射が内耳バリアもラットの聴覚障害も引き起こしませんでした。以前に実証された重要な炎症性生物学的マーカーを分析することにより[11、22]、LPNおよびLPN + Gd-DOTAは蝸牛に炎症反応を誘発しませんでした。正円窓膜は評価されませんでしたが、本研究では卵円窓経路がLPNの正円窓アプローチよりも優れていることが示されたため、アブミ骨と卵円窓に炎症がないことから、正円窓膜に明らかな炎症反応が見られませんでした。
ガドリニウムキレートの静脈内注射後の内耳MRIは、ミトコンドリア毒素によって誘発される血液外リンパおよび血液内リンパバリアの酸化ストレスを介した破壊を検出することができます[23]。 AgNPは、活性酸素種(ROS)の生成とJunアミノ末端キナーゼ(JNK)の活性化を通じて細胞障害を引き起こし、シトクロムCのサイトゾルへの放出と、Baxのミトコンドリアへの移行を引き起こすことが報告されています[24 ]。鼓室内注射では、AgNPが内耳に入り、ラット内耳の生物学的障壁に透過性の変化を誘発しました[11、25]。 LPNは、鼓室内注射後にサイズ依存パターンでラット内耳にも侵入し、直径95 nmのLPNは、中内耳バリアを通過する際に最も高い効果を示しました[3]。今回の調査では、LPNの平均サイズは100〜115 nmであり、最も効率的なサイズよりもわずかに大きかった。 LPN + Gd-DOTAは、このサイズのLPNが内耳に入ったことを示しました。これは、以前のレポート[3]と一致しています。ただし、内耳へのLPNの侵入は、血液外リンパおよび血液内リンパバリアの透過性の変化を引き起こしませんでした。この結果は、LPNが内耳にとって安全であることを示唆しています。正常な聴覚機能を示すABRの結果は、MRIの結果を裏付けました。
腎虚血性再灌流傷害において、ヒアルロン酸分泌と透過性変化および微小循環炎症との間に関連があった[12]。以前の研究はまた、AgNPがラットの蝸牛にヒアルロン酸の蓄積を引き起こしたことを示しました[11]。 CD44とトール様受容体2/4(TLR2 / 4)は、ヒアルロン酸の受容体として機能し、生物学的反応を引き起こします[13、14]。 CD44はまた、炎症部位へのT細胞の動員と、T細胞を介した内皮損傷の調節に加えて、細胞の取り込みと分解を通じてヒアルロン酸の代謝を仲介します[18]。本研究では、ヒアルロン酸、CD44、およびTLR2がラット蝸牛で検出されました。 LPN + Gd-DOTAは、らせん靭帯線維細胞におけるヒアルロン酸の分泌を減少させました。 CD44とTLR2の発現は、LPNまたはLPN + Gd-DOTAのいずれかの鼓室内注射後に変化しませんでした。蝸牛にヒアルロン酸を含む総グリコサミノグリカンは、LPNおよびLPN + Gd-DOTAの投与による影響を受けませんでした。ラット蝸牛のヒアルロン酸分布に対するLPNの影響は、透過性の変化も難聴も引き起こさず、修飾が無害であることを示しています。マクロファージは、低分子量ヒアルロン酸の炎症誘発性反応または高分子量ヒアルロン酸の抗炎症反応のいずれかに対応する、ヒアルロン酸の分子量に依存する表現型の変化を受けることが報告された[26]。蝸牛で観察されたヒアルロン酸分布の小さな変化は、高分子量と抗炎症機能を持っている可能性があります。したがって、ラットの蝸牛に炎症反応の兆候はありませんでした。
あぶみ骨のフットプレート細胞で観察されたアポトーシスは、正常な生物活性である可能性があります。耳硬化症を不活化するために必要に応じて、あぶみ骨足板細胞の生存とアポトーシスのバランスが報告されました[27]。 LPN + Gd-DOTAの投与は、ラットのあぶみ骨のアポトーシスに影響を与えませんでした。
結論
本研究は、リポソームナノキャリアの鼓室内注射が内耳の生物学的障壁を損なうことも、ラットの難聴を引き起こすこともないことを示した。重要な炎症メカニズムは、リポソームナノキャリアの投与によっても活性化されなかった。結果は、リポソームナノキャリアの鼓室内注射がラットの蝸牛にとって安全であることを示唆しました。
メソッド
資料
スフィンゴシン(Sph)、1-ステアロイル-2-オレオイル-sn-グリセロ-3-ホスホコリン(SOPC)、および1,2-ジステアロイル-sn-グリセロ-3-ホスホエタノールアミン-N- [メトキシ(ポリエチレングリコール)-2000](アンモニウム塩)[DSPE-PEG-2000]は、Avanti極性脂質(Alabaster、USA)から購入しました。 DiI(Vybrant DiI細胞標識溶液、溶媒中1 mM)およびN-(6-テトラメチルローダミンチオカルバモイル)-1,2-ジヘキサデカノイル-sn-グリセロ-3-ホスフォエタノールアミン、トリエチルアンモニウム塩(TRITC-DHPE)はThermo FisherScientificから購入しました。 (ウォルサム、米国)。 Gd-DOTA(DOTAREM)は、フランスのCedexのGuerbetから来ました。 HepesとEDTAはSigmaからのものでした。脂質の純度は、クロロホルム/メタノール/水の混合物(65:25:4、v / v / v)で開発したケイ酸コーティングプレート(Merck、ダルムシュタット、ドイツ)での薄層クロマトグラフィーを使用して評価しました。ヨウ素染色後、および適切な場合はUV照射時にプレートを検査したところ、不純物は検出されませんでした。脂質濃度は、SuperG(Kibron、Espoo、フィンランド)を使用して重量分析で決定されました。高精度のマイクロバランス。ポリビニルピロリドンで安定化されたAgNPは、Colorobbia(Firenze、Italy)から提供されました。 AgNPは脱イオン水(370.7 mM)に分散されており、走査型電子顕微鏡により、AgNPは粒子サイズが約100nmの球状体であることがわかりました。動的光散乱(DLS)は、117±24nmの平均流体力学的サイズと-20±9mVの平均ゼータ電位を示しました。
内耳へのナノキャリアの取り込みの視覚化と生物学的バリア機能の評価では、体重334〜348gの5匹のオスのSpragueDawleyラットが、フィンランドのヘルシンキ大学の実験動物センターであるBiomedicum Helsinkiから提供されました(これはBiomedicumでのMRI実験のために動物を提供する定義された動物センター。 ABRおよび組織学的研究では、体重300〜400gの18匹のSpragueDawleyラットが、フィンランドのタンペレ大学医学部の実験動物ユニットから提供されました。各研究における動物の割り当てを表2に示しました。すべての動物実験は、タンペレ大学の倫理委員会によって承認されました(許可番号:ESAVI / 3033 / 04.10.03 / 2011)。動物の世話と実験手順は、ヨーロッパの法律に従って実施されました。 Gd-MRI研究の動物は、誘導のために5%のイソフルランと酸素の混合物(流量1.0 L / min)、フェイスマスクを介した維持のために3%のイソフルランで麻酔されました。 ABRおよび組織学的研究のための動物は、0.5 mg / kgの塩酸メデトミジン(Domitor ® )の混合物で麻酔されました。 、Orion、エスポー、フィンランド)および75 mg / kgの塩酸ケタミン(Ketalar ® 、ファイザー、ヘルシンキ、フィンランド)腹腔内注射とそれに続くエンロフロキサシン(Baytril ® の筋肉内注射) 獣医、オリオン、トゥルク、フィンランド)潜在的な感染を防ぐために、10 mg / kgの用量で。動物の目はViscotears®(Novartis Healthcare A / S、コペンハーゲン、デンマーク)によって保護されていました。
<図>Gdを使用した場合と使用しない場合のLPNの準備と特性評価
Gdを含むLPNの準備
見かけの流体力学的粒子径( Z )を持つ単層ベシクルのLPN av )Gd-DOTAを含む110±15 nmは、以前に公開された方法[6]に従って調製されました。 1 mM Gd-DOTA含有LPN(LPN + Gd-DOTA)の濃度は、500 mmol / LのGd-DOTAをカプセル化する1mMリポソームを指します。
空白のLPNの準備
Z の単層ベシクルのブランクLPN av 115±10nmは以前の出版物[6]に従って調製されました。
LPNの管理
5%イソフルラン-酸素混合物(流量1.0 L / min)を含むイソフルランによる全身麻酔下で、手術顕微鏡下で鼓膜貫通部から50μlのLPNまたはLPN + Gd-DOTAを左中耳腔に注入しました。 (OPMI1-F、Carl Zeiss、イエナ、ドイツ)。同量の脱イオン水(H 2 O)陰性群に割り当てられたラットに経鼓膜注射した。注射後、さらに測定する前に、注射した耳を上に向けて15分間、動物を横向きに保ちました。
Gd-MRIを使用した生物学的バリア機能の評価
LPN + Gd-DOTAの鼓室内注射を受けた1匹の動物を選択して、造影剤なしのMRIを使用して内耳のLPNの分布を示しました。ブランクLPNの鼓室内注射を受けた2匹の動物は、生物学的バリア機能の評価のためにMRI研究に従事した。 AgNP(370.7 mM、40μl)の鼓室内注射を受けた2匹の動物を陽性対照として使用しました。すべての研究において、注射を行わない反対側の耳を陰性対照として使用した。ボア径155mmの4.7TMRスキャナー(PharmaScan、Bruker BioSpin、Ettlingen、ドイツ)を使用しました。最大勾配強度は300mT / mで、立ち上がり時間は80μsでした。ガドリニウム-テトラアザシクロドデカン-四酢酸(Gd-DOTA、500 mM、DOTAREM、Guerbet、Cedex、フランス)溶液を、MRI測定の2時間前に尾静脈(0.725 mM / kg)に注入しました。イメージングプロトコルとリラクゼーションエンハンスメント(RARE)シーケンスによる迅速な取得は、以前の出版物[10]に従って適用されました。 MRIスキャンは、鼓室内注射後のいくつかの時点で開始されました。約5時間の最初のMRI時間は、リポソームナノ粒子が中耳から内耳に浸透する時間を基準として決定されました[1、3、6]。 8日間の最終イメージング時間は、潜在的な急性炎症の経過とスキャナーの可用性に応じて選択されました。 ParaVision PV 4.0(Bruker、MA、USA)ソフトウェアは、MR画像の後処理と定量化に使用されました。
ABR測定
ブランクLPNとGd含有LPNの両方の注射を受けた動物の聴覚機能は、カスタムメイドの防音室でBioSig32(Tucker Davis Technologies、FL、USA)を使用したABR測定を使用して評価されました。クリックおよびトーンバースト刺激時のABRしきい値は、LPNの投与前および投与後の特定の時点で記録されました。最初のABR測定は、AgNPの投与後2日後に行われ、動物が注射中に全身麻酔から回復し、注射された溶液が中耳腔から完全に除去されることを保証しました。注射後4日間の2回目のフォローアップ時間は、ミトコンドリア障害によって誘発される蝸牛の細胞死のピーク時間に近いために選択されました[22]。 7日間の3番目のフォローアップ時間は、ミトコンドリア毒素誘発性難聴の動物モデルで一時的なしきい値のシフトが大幅に承認されたままであった時点です[28]。 ABRの記録手順は、以前のレポート[11]に従いました。
LPN投与後のラット蝸牛におけるグリコサミノグリカン染色
潜在的な炎症性浸潤を評価するためのヘマトキシリン-エオシン染色と、LPNの投与後の蝸牛における潜在的なグリコサミノグリカンの蓄積を評価するための過ヨウ素酸シッフ染色は、7日間にわたるABR測定後に以前の出版物に従って実施されました[11]スライスが観察され、デジタル画像が光学顕微鏡(オリンパスDP25カメラを搭載したライカDM2000顕微鏡)で取得し、さらに分析します。
ヒアルロン酸と受容体の蛍光抗体染色
ヒアルロン酸、CD44、およびTLR2の免疫蛍光染色は、7日間にわたるABR測定後の以前の出版物に従って実施されました[11、21]。
細胞死の検出
報告された手順[11]に従って、末端トランスフェラーゼ(TdT)を使用して、アポトーシス細胞のDNA鎖の遊離3'OH切断をTMR-dUTP(TUNEL染色)で標識することにより、蝸牛における潜在的な核DNA断片化を調査しました。組換えDNaseI(Fermentas、Vantaa、フィンランド、50 mM Tris / HCl中100U / ml、pH 7.5、1 mg / mlウシ血清アルブミン)に37°Cで10分間曝露したスライスは、前にDNA鎖切断を誘発しました標識手順は、陽性対照として利用された。サンプルは共焦点顕微鏡で観察されました。
共焦点顕微鏡
サンプルは、Andor iQ 2.8ソフトウェア(Andor Technology、Belfast、UK)がインストールされたAndor共焦点システムと組み合わせたNikon倒立顕微鏡(ECLIPSE Ti)で観察されました。励起レーザーは、Andorレーザーコンバイナーシステムからの488 nm(緑色励起)および568 nm(赤色励起)であり、対応する発光フィルターは525/50(Alexa Fluor-488)および607/45 nm(Cy TM)でした。 3およびTMRレッド)。 DAPIは、発光ダイオードから生成された405 nmの光で励起され、450〜465nmのフィルターを使用して検出されました。
分析と統計
ImageJ(1.45S、米国国立衛生研究所、ベセスダ、米国)ソフトウェアを信号強度測定に使用しました。過ヨウ素酸シッフ染色の光学顕微鏡検査では、らせん靭帯、らせん靭帯、血管条などの関心領域(ROI)をフリーハンド選択ボタンを使用して選択しました。 「測定」関数を使用して、ROIの平均グレースケール値を取得しました。これは、染色強度と逆相関していました。蛍光抗体染色の共焦点顕微鏡検査では、線条体基底細胞、らせん靭帯細胞、らせん靭帯、らせん靭帯などのROIを、フォトショップCS6(バージョン13.0、Adobe Systems Software Ireland Ltd、ダブリン、アイルランド)プログラムを使用して抽出し、ImageJプログラムにインポートしました。 。画像は個々のチャネルに分割され、さらに定量化するために緑(CD44およびTLR2に対応)および赤(ヒアルロン酸に対応)のチャネルが選択されました。 「しきい値」は「画像」メニューの「設定」ボタンを使用して調整し、「しきい値に制限」オプションを選択し、「分析」メニューの対応するチャネルに「直接」を定義する必要があります。次に、同じメニューの「測定」機能を使用して、染色強度と逆相関するグレースケール値を取得しました。
統計分析は、IBM ® を使用して実行されました。 SPSS ® Statisticsバージョン20ソフトウェアパッケージ(SPSS Inc.、シカゴ、米国)。一元配置分散分析とクラスカル・ウォリス検定を使用して、グリコサミノグリカンとヒアルロン酸分泌物の染色のABRしきい値シフトと信号強度(グレースケール)、およびLPN注入耳グループと生理食塩水注入耳グループ間のTLR2とCD44染色を比較しました。さまざまなターンの間の異なる蝸牛構造。事後分析として、最小有意差(LSD)テストを使用しました。グレースケール分析の数値が高いほど、染色の信号強度が低くなります。 p <0.05は統計的に有意であると認められました。
略語
- ABR:
-
聴性脳幹反応
- AgNPs:
-
銀ナノ粒子
- dH 2 O:
-
脱イオン水
- GAB1:
-
成長因子受容体結合タンパク質2関連結合タンパク質1
- Gd-DOTA:
-
ガドリニウム-テトラ-アザシクロ-ドデカン-テトラ酢酸(DOTAREM)
- Gd-MRI:
-
ガドリニウム造影磁気共鳴画像法
- JNK:
-
Junアミノ末端キナーゼ
- LPN + Gd-DOTA:
-
Gd-DOTAを含むLPN
- LPN:
-
リポソームナノキャリア
- ROI:
-
関心領域
- ROS:
-
活性酸素種
- TLR:
-
Toll様受容体
ナノマテリアル