


多層カーボンナノチューブでトリガーされたトマトの側根形成の根底にある新しいメカニズム:一酸化窒素の関与
要約
豊富な研究により、多層カーボンナノチューブ(MWCNT)は植物に有毒であることが明らかになりました。ただし、MWCNTがさまざまな環境の手がかりへの根系の適応性の重要な要素である側根(LR)形成に影響を与えるかどうか、またはどのように影響するかについては、依然として議論の余地があります。このレポートでは、MWCNTがトマトの苗の根に侵入する可能性があることを発見しました。 MWCNTの投与は、ほぼ用量依存的にトマトのLR形成を促進しました。内因性一酸化窒素(NO)の生成は、MWCNTによってトリガーされ、Greiss試薬法、電子常磁性共鳴(EPR)、レーザー走査型共焦点顕微鏡(LSCM)、およびNOのスカベンジャーによって確認されました。 MWCNTによって引き起こされるNO合成とLR形成は、そのスカベンジャーによる内因性NOの除去によって明らかにブロックされたため、LR発生の誘導においてMWCNTとNOの間に因果関係が存在します。 NO生成酵素硝酸レダクターゼ(NR)の活性は、MWCNTに応答して増加しました。 NRのタングステン酸塩阻害は、NO産生を損なうだけでなく、MWCNTによって引き起こされるLR形成を無効にしました。 N の追加 G -哺乳類の一酸化窒素シンターゼ(NOS)様酵素の阻害剤であるニトロ-1-アルギニンメチルエステル(l-NAME)は、LR形成に影響を与えませんでした。まとめると、NOは、少なくとも部分的にNRを介して、LR発生のMWCNT制御において下流のシグナル伝達分子として機能する可能性があることを提案しました。
はじめに
カーボンナノチューブには多くの生物学的および生物医学的用途があります[1、2]。細胞膜に容易に浸透する独自の能力により、カーボンナノチューブのバイオセーフティは常に議論の的となっています[3、4]。一方、カーボンナノチューブの生産と使用は急速に成長しているため、人間と哺乳類、そして最近では植物におけるその細胞毒性の詳細なメカニズムを特徴づけることが重要になります[3,4,5,6,7,8,9]。植物とそのコミュニティは人間と環境にとって非常に重要であり、バイオソリッド肥料の施用による土壌への蓄積により、カーボンナノチューブへの曝露のリスクがあることはよく知られています[6、10、11]。カーボンナノチューブの重要なメンバーとして、グラフェンの複数の圧延層からなる多層カーボンナノチューブ(MWCNT)の毒性が広く研究されてきました。哺乳類での研究により、MWCNTと単層カーボンナノチューブの両方への曝露が、ヒトケラチノサイトとA549細胞に酸化的損傷とNF-κB活性化を誘発することが明らかになりました[9、12]。 MWCNTと単層カーボンナノチューブは原形質膜と融合する可能性があるため、脂質過酸化と酸化ストレスによる細胞損傷を引き起こします[9、11、13、14]。 MWCNTによって引き起こされる細胞毒性と酸化ストレス、および中程度の炎症反応が、ヒト臍帯静脈内皮細胞で観察されました[15]。以前の研究では、赤ホウレンソウにおけるMWCNTの主な毒性は、主に活性酸素種(ROS)の過剰産生に由来し、毒性効果はアスコルビン酸の補給によって逆転する可能性があることが示唆されました[7]。この意味で、MWCNTは、動物または植物のいずれかにおいて、生物にとって新たなストレス要因と見なされています。
根の構造の重要な決定要因である側根(LR)の形成は、さまざまなストレスに対する適応応答の指標と見なされてきました[16]。高等植物では、LRの形成は、植物ホルモンと、水の利用可能性、栄養素、および低酸素症や重金属ストレスなどの非生物的ストレスを含む幅広い環境的手がかりの影響を受けます[17、18、19]。一方、十分な証拠により、LRの形成は物理的なサポートとして機能するだけでなく、植物の成長と発達のための水分と栄養素の取り込みにも寄与することが確認されています[19、20、21]。さまざまな環境の手がかりが、LR形成の促進や根の伸長の阻害など、いくつかの特定のストレス誘発性形態形成反応(SIMR)表現型を引き起こす可能性があります[17]。LR形成の調節もオーキシンなどの植物ホルモンによって厳密に制御されています。そして、オーキシンに応答した細胞周期調節遺伝子の活性化が示唆されました[19、22]。一方、オーキシンによって引き起こされる根の器官形成へのいくつかの小分子の関与は、キュウリ、トマト、大豆、および菜種植物で確認されました[23、24、25、26、27]。これらの小分子には、過酸化水素(H 2 O 2 )、一酸化窒素(NO)、一酸化炭素(CO)、および水素ガス(H 2 。
これらの中で、フリーラジカルガスであるNOは、植物において複数の生理学的機能を持っていることが示されています[28、29]。ストレスに対する植物の適応の強化に加えて、NOの機能には、根毛の発達、不定の発根、および側根形成の促進が含まれます[30、31、32、33]。前述のプロセスはとらえどころのないままです。動物では、l-アルギニンからのNOの合成は、ヘム含有酵素である一酸化窒素合成酵素(NOS)によって触媒されます[34]。 NOS酵素をコードする遺伝子は植物では同定されていませんが、哺乳類のNOS様活性は広く検出されており[35、36]、 N などの哺乳類のNOSの阻害剤です。 G -ニトロ-1-アルギニンメチルエステル塩酸塩(l-NAME)は、植物のNO生成を阻害する可能性があります[25、33、36、37、38、39]。重要なことに、十分な遺伝的証拠により、植物の窒素代謝に関与するよく知られた酵素である硝酸レダクターゼ(NR)によってNOが生成されることが明らかになりました[28]。気孔閉鎖および低温順化におけるNR媒介NO産生の関与は遺伝的に実証されている[37、38]。私たちの以前の研究は、NR依存性のNO合成がオーキシン誘発性の水素ガス媒介性の側根形成に関与していることを示しました[39]。
これまで、MWCNT [40,41,42,43]、金ナノ粒子(Au NP、[44])、酸化亜鉛などのナノ材料を補充すると、LRの形成、促進、または阻害におけるさまざまな応答が、さまざまな植物種でそれぞれ報告されました。ナノ粒子(ZnO NP [45、46];)、二酸化チタンナノ粒子(TiO 2 NP [46];)、および酸化グラフェン(GO [47,48,49];)(表1)であり、上記の応答におけるNOの役割の明確な証拠を提供した研究はまだありません。この研究では、Greiss試薬法、レーザー走査型共焦点顕微鏡(LSCM)、および電子常磁性共鳴(EPR)分析による内因性NOの検出により、MWCNT処理トマト苗でNOレベルが増加したことが明らかになりました。その後、LR形成が観察された。さらに、NRおよび哺乳類のようなNOS活性を阻害するNOスカベンジャーおよびアンタゴニストを使用して内因性NOレベルを操作することにより、MWCNTによって引き起こされるLR形成におけるNOの関与を研究します。さらなる実験により、NR依存性NOは、少なくとも部分的に、MWCNTに応答したLR形成に不可欠である可能性があることが明らかになりました。したがって、この研究は、植物におけるナノマテリアルの生物学的影響を理解するための新しいウィンドウを開きます。
<図>材料と方法
化学薬品
特に明記されていない限り、他のすべての化学物質はSigma-Aldrich(セントルイス、ミズーリ州、米国)から入手しました。 Sigma-Aldrichから購入したMWCNTは、前述のように特性評価されました[50]。 MWCNTの外径、内径、および長さは、それぞれ6–12 nm、2.5–5 nm、および1–9μmでした。超音波処理後、得られたホモジネートコロイド懸濁液を滅菌して使用しました。
その他のカーボンナノ粒子は、単層カーボンナノチューブ(SWCNT、XFS22、純度> 95%、直径1〜2 nm、長さ5〜30μm、特殊表面積> 1075 m)を含め、南京XFNANO Materials Tech Co.、Ltd。から入手しました。 2 / g)、グラフェン(XF001W;純度〜99%、直径0.5–5μm、厚さ〜0.8 nm、単層比〜80%、BET表面積500〜1000 m 2 / g;電気抵抗率≦0.30Ω.cm)、および活性炭(AC、XFP06;純度> 95%、粒子サイズ5±1μm、細孔容積1–1.2 cm 3 / g、アパーチャ2.0–2.2 nm、特殊表面積〜1500–1700 m 2 /g)。
さらに、ニトロプルシドナトリウム(SNP)がNO放出化合物として使用されました[30、31、32、33]。 2-(4-カルボキシフェニル)-4,4,5,5-テトラメチルイミダゾリン-1-オキシル-3-オキシドカリウム塩(cPTIO)は、NOのスカベンジャーと見なされていました[51,52,53,54]。タングステン酸塩(Tg; NRの阻害剤[28、33、37、55,56,57];)および N G -ニトロ-L-アルギニンメチルエステル塩酸塩(NAME;哺乳類のNOS様酵素の阻害剤[25、33、36、37、38、39];)も適用されました。この研究では、上記の化学物質の濃度がパイロット実験で決定され、そこから有意な反応が観察されました。
植物の材料と成長条件およびLR形成の決定
トマト( Solanum lycopersicum L.)種子「江蘇14」は、中国江蘇省南京市の江蘇農業研究所から提供されました。同じサイズの選択された種子を、25±1°Cの蒸留水中で暗所で3日間発芽させました。次に、幼根が2〜3mmの選択された同一の苗木を、示された濃度のMWCNT、200 nM 1-ナフチル酢酸(NAA;よく知られているオーキシン)、0.1 mM SNP、0.2 mM cPTIO、20を含む6mL処理溶液に移しました。 μMタングステン酸塩(Tg)、0.2 mM NAME、および5 mg / mL単層カーボンナノチューブ(SWCNT)、グラフェン、および活性炭素(AC)を含むその他のカーボンナノ粒子を、指定された時点で単独または組み合わせて使用します。苗木は、200μmolm -2 の光強度の照明インキュベーター(25±1°C)で育てられました。 s -1 14/10 h(明/暗)日長で。
処理後、写真を撮り、Image Jソフトウェア(http://rsb.info.nih.gov/ij/)を使用して苗木あたりの出根(> 1 mm)の数と長さを決定しました[39 、58]。前に説明したように、その後の分析には側根誘導性セグメントのみを使用しました。
透過型電子顕微鏡によるMWCNT分布のイメージング
トマト実生の根におけるMWCNTの分布は、透過型電子顕微鏡(TEM; JEOL、JEM-200CX、東京、日本)を使用して特徴づけられました。 TEM分析のサンプル準備は、以前のプロトコル[59]に従った。
レーザー走査型共焦点顕微鏡による内因性NOのイメージング
NOイメージングは、かなり特異的なNO蛍光プローブ4-アミノ-5-メチルアミノ-2 '、7'-ジフルオロフルオレセインジアセテート(DAF-FM DA)を使用して実施しました。プローブを完全に洗浄した後、Zeiss LSM 710共焦点顕微鏡(Carl Zeiss、ドイツ、オーバーコッヘン、励起は488 nm、発光は500〜530 nm、NO分析)を使用して画像を取得しました。私たちの実験では、20の個別のサンプルがランダムに選択され、処理ごとに測定されました。写真は同じ結果を表しています。
グリース試薬アッセイによって含有量が決定されていない
以前に記載された方法[50]に従って、Griess試薬アッセイでNO含有量を測定しました。重要なのは、植物の硝酸塩と亜硝酸塩の濃縮含有量によって引き起こされる干渉を回避するために、200μMのcPTIO(NOのスカベンジャー)で30分間プレインキュベートした同一のサンプルをブランクサンプルと見なしたことです。 Griess試薬を30分間添加した後、540 nmで吸光度を記録し、NaNO 2 の標準曲線と比較してNO含有量を測定しました。 。
電子常磁性共鳴(EPR)によるNOの決定
以前の方法[39、55、60]に従って、電子常磁性共鳴(EPR)を使用したNOレベルの決定が実行されました。有機溶媒層を使用して、Bruker A300分光計(Bruker Instrument、カールスルーエ、ドイツ)で次の条件下でNOを測定しました。マイクロ波周波数、9.85 GHz;マイクロ波電力、63.49 mW;および変調周波数、100.00kHz。
硝酸レダクターゼ(NR)活性の測定
NR活性は、以前の方法[57]に従って、540nmで分光光度的に検出されました。生成された亜硝酸塩は、3 MHCl中の1%(w / v)スルフェニルアミド1 mLと0.02%(v / v)n-(1-ナフチル)-エチレンジアミン1 mLを添加することにより、540nmで分光光度的に測定されました。
統計分析
示されている場合、結果は、それぞれに3つの生物学的複製を用いた3つの独立した実験の平均値±SEとして表されました。統計分析は、SPSS Statistics17.0ソフトウェアを使用して実行されました。統計分析の場合、ダンカンの複数のテスト( p <0.05)が適切に選択されました。
結果
MWCNTは根細胞への侵入だけでなく、LR形成を促進します
LRの形成は、ルートシステムアーキテクチャの主要な決定要因です。 LR形成に対するMWCNTの影響を調査するために、3日齢のトマト実生をさまざまな濃度のMWCNT(0.05、0.5、5、および50 mg / mL)と3日間インキュベートしました。 1-ナフチル酢酸(NAA)の適用は陽性対照と見なされた。私たちの実験では、LR数と長さの両方がLR形成の2つのパラメーターとして決定されました。図1に示すように、対照サンプルと比較して、外因性MWCNTは、ほぼ用量依存的にLR形成を有意に誘導し、5および50 mg / mLで最大の効果を示しました。 200 nMのNAAを投与した場合にも、同様の誘導反応が観察されました。 MWCNTのコストとLR形成における誘導応答を考慮して、5 mg / mLのMWCNTを以下の実験に適用しました。
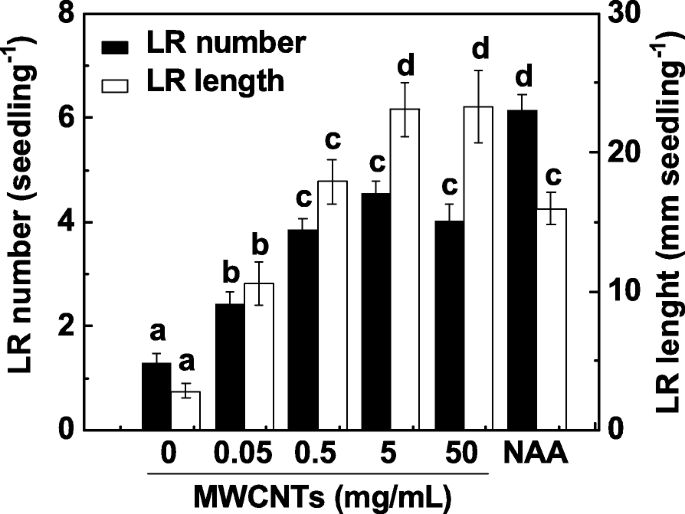
MWCNTによって誘発されたトマトのLR形成は、ほぼ用量依存的に行われた。 3日齢のトマト実生をそれぞれ200nMのNAAと示された濃度のMWCNTで処理しました。次に、実生あたりの出芽した側根(> 1 mm)の数と長さを、3日間の処理後に決定しました。 3つの生物学的複製に30(10×3)の植物があり、実験は3回行われた。データは平均値±SEです。実験の各セット内で、同じ文字で示されたバーは p で有意差はありませんでした <ダンカンの複数のテストによる0.05レベル
LR形成の誘導におけるMWCNTの特定の機能を検証するために、MWCNTの他の同素体にもそのような誘導効果があるかどうかをさらに調査します。図2aに示すように、これらのカーボンナノ材料はすべて、シュートの成長に対して毒性作用を示しました(データは示していません)。興味深いことに、MWCNT、単層カーボンナノチューブ(SWCNT)、グラフェン、および同じ濃度(5 mg / mL)の活性炭を適用すると、化学物質を含まない対照植物と比較して、LRの数と長さが異なる結果になる可能性があります。 (図2b)。これらの化学物質の中で、MWCNTで培養したトマトの苗で最大の誘導反応が発見されました。
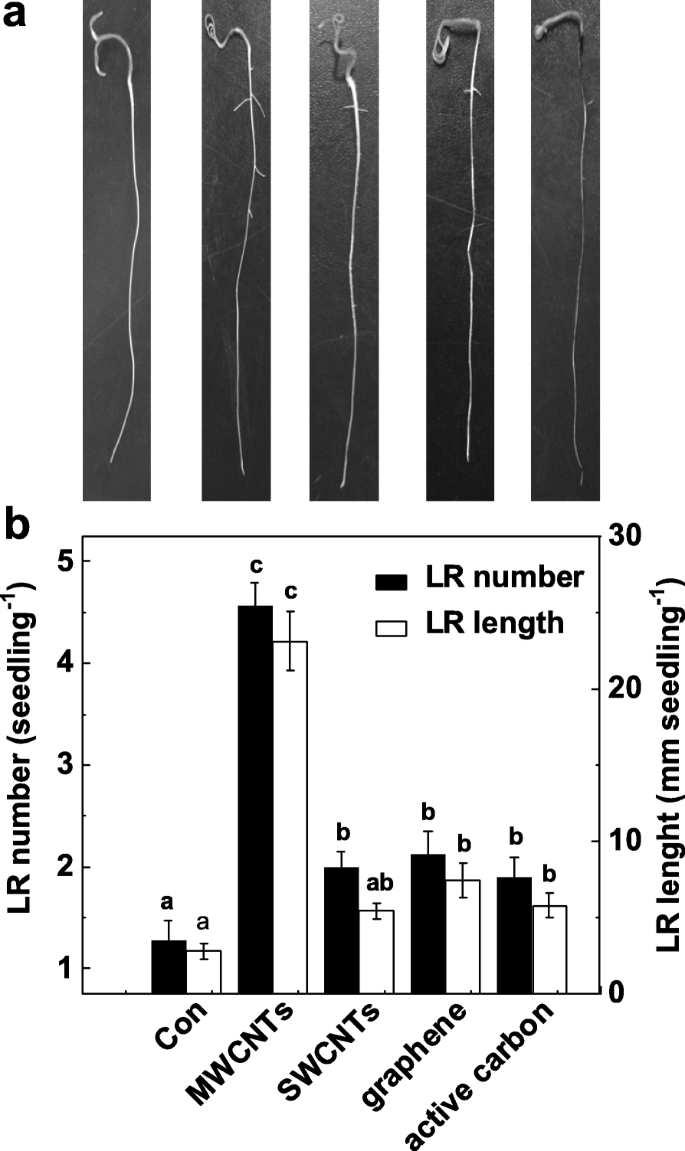
異なるカーボンナノ粒子に応じた側根形成の変化。 3日齢のトマトの苗木を、蒸留水(Con)、5 mg / mL MWCNT、単層カーボンナノチューブ(SWCNT)、グラフェン、および活性炭(AC)でそれぞれさらに3日間処理しました。 a その後、代表的な写真を撮りました。 b 次に、実生あたりの出芽した側根(> 1 mm)の数と長さも決定しました。スケールバー=50mm。 3つの生物学的複製に30(10×3)の植物があり、実験は3回行われた。データは平均値±SEです。実験の各セット内で、同じ文字で示されたバーは p で有意差はありませんでした <ダンカンの複数のテストによる0.05レベル
透過型電子顕微鏡(TEM)の助けを借りて、MWCNTの分布を簡単に評価することができます。図3に示す結果は、MWCNTを外因的に適用すると、トマトの実生に吸収され、MWCNTの分布が根の細胞にあることを示しています。苗の根はMWCNTを含む溶液中で直接培養されるため、この結果は理解できます。
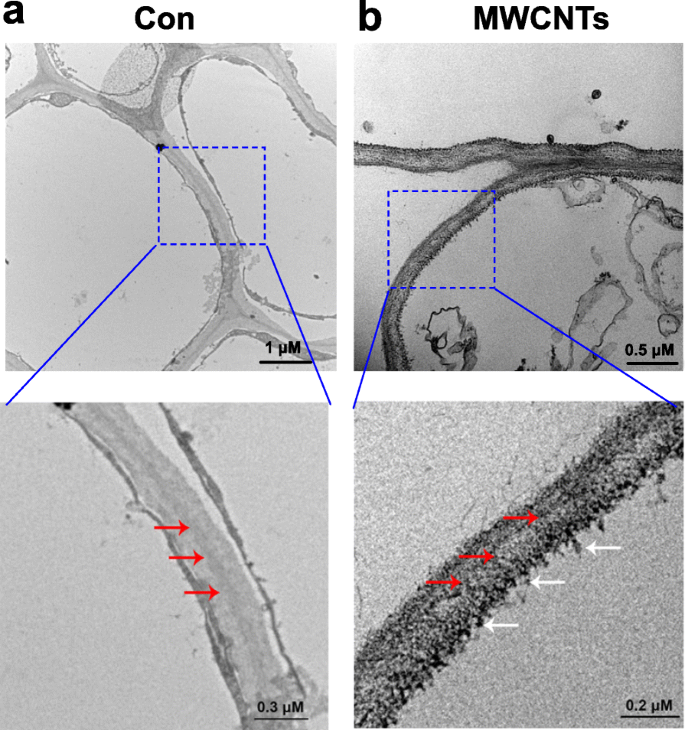
トマトの根におけるMWCNTの分布。蒸留水で処理した3日齢のトマト苗のTEM画像(Con; a )または5 mg / mLMWCNT( b )1日かかりました。赤い矢印は細胞壁を示し、白い矢印はMWCNTを示します
MWCNTによって誘発されたNO合成とその後のLR形成は、NOのスカベンジャーであるcPTIOに敏感でした
NOがMWCNTによって誘発されるLR形成にも関与しているかどうかを調べるために、MWCNTによって誘発されるLR形成におけるNOの機能を、NO放出化合物とスカベンジャーを使用して内因性NOレベルを操作することによって評価しました。以前の結果[31]と同様に、ニトロプルシドナトリウム(SNP)の投与は、LR形成の誘導をもたらす可能性があり、SNPとMWCNTを一緒に適用した場合に相加的応答が観察されました(図4)。 2-(4-カルボキシフェニル)-4,4,5,5-テトラメチルイミダゾリン-1-オキシル-3-オキシドカリウム塩(cPTIO; NOのスカベンジャー)を添加した場合、MWCNTによって引き起こされるLR形成の促進反応は有意でした。障害者。単独で、cPTIOは化学物質を含まない対照と比較してLRの発達を阻害する可能性があり、根の器官形成における内因性NOの重要な役割を示しています。
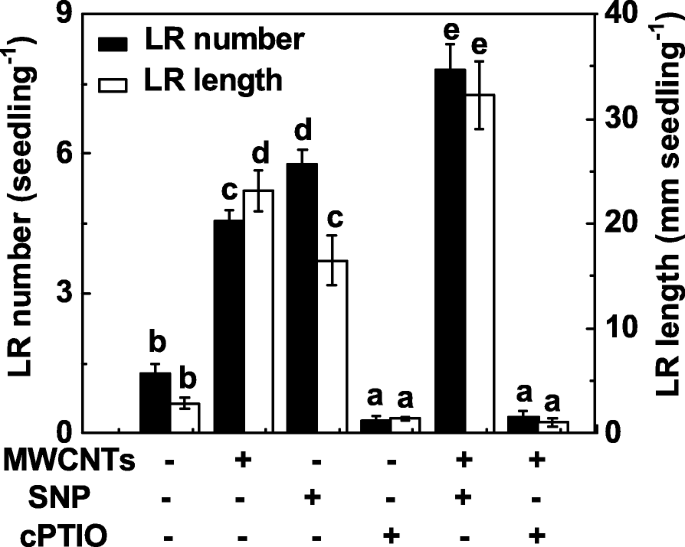
MWCNTによって誘発されたLR形成は、そのスカベンジャーであるcPTIOによる内因性NOの除去に敏感でした。 3日齢のトマト苗を蒸留水、5 mg / mL MWCNT、0.1 mM SNP、0.2 mM cPTIOで、単独または組み合わせて3日間処理しました。その後、苗木ごとに出現した側根の数と長さ(> 1 mm)を決定しました。 3つの生物学的複製に30(10×3)の植物があり、実験は3回行われた。データは平均値±SEです。実験の各セット内で、同じ文字で示されたバーは p で有意差はありませんでした <ダンカンの複数のテストによる0.05レベル
MWCNT応答における内因性NOの重要な役割をさらに評価するために、invivoでのNO産生の時間経過を最初にGreiss試薬法で検出しました。上記の測定中、cPTIOで前処理された同一のろ液は、正確な結果を得るためにブランクと見なされました。トマト実生の根におけるNO産生は、MWCNT処理後24時間まで劇的に増加し、その後初期レベル(48時間;図5a)に回復することが観察されました。 MWCNTによって24時間トリガーされた内因性NOの最大レベルを超えると、NOのスカベンジャーであるcPTIOによって明らかに廃止され、NOの特定の役割が示唆されました。
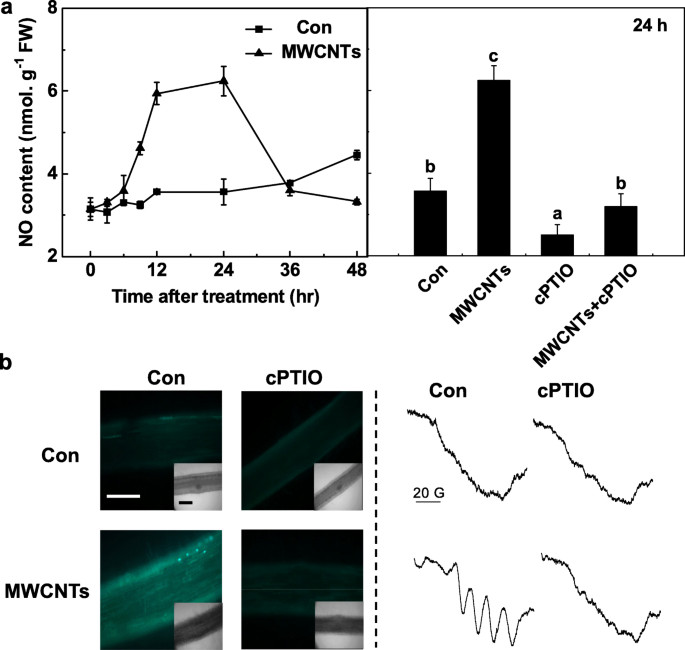
MWCNTによって誘発されたNO生成は、NOのスカベンジャーであるcPTIOによってブロックされました。 3日齢のトマトの実生を蒸留水と5mg / mLのMWCNTで、それぞれ0.2mMのcPTIOの有無にかかわらず処理しました。 a グレイス試薬法を使用して決定されたNR活性の変化(左)、およびNO産生(右)。 b 24時間の処理後、NO信号をLSCM(左)とEPR(右)で分析しました。スケールバー=0.1mm。データは平均値±SEです。同じ文字で示されたバーは、 p で大きな違いはありませんでした
上記の結果を確認するために、LSCMとESRの両方を採用しました。まず、トマトの実生の根における内因性NOレベルの変化を、細胞透過性でかなりNO特異的な蛍光プローブDAF-FM DAを使用してNOを標識し、LSCMでイメージングすることによって監視しました。以前の結果(図5a)と同様に、cPTIOの存在下では、MWCNTによって引き起こされるDAF-FM依存性の蛍光強度の増加は大幅に廃止されました(図5b)。これらの結果は、DAF-FMによってトリガーされる蛍光がトマトの実生の根の内因性NOレベルに関連していることを意味します。
MWCNTによって誘発されたNO生成は、EPR分光法によって確認されました。予想通り、MWCNTで24時間処理した実生の根は、NO複合体の典型的な超微細構造のトリプレットを示しました。ただし、cPTIOの追加は上記の信号を廃止し、MWCNTへの曝露が強いNO生成をもたらしたことを示しています(図5b)。まとめると、これらのデータは、トマトの実生におけるMWCNTによって引き起こされるLR形成には合成が必要ない可能性があることを示唆しています。
NRはMWCNTによるNO産生とその後のLR形成に関与する可能性があります
NRと哺乳類様NOSは植物のNO合成に関連する2つの主要な酵素であるため、その後の実験ではタングステン酸塩(NR阻害剤)とNAME(哺乳類NOS阻害剤)の両方を適用しました。ここで、タングステン酸塩処理は、MWCNT処理したトマト実生の根におけるLR形成の促進を実質的に抑制しました(図6)。比較すると、MWCNTによって引き起こされるLR形成の誘導は、NAMEの追加によって強く阻害されなかった。これは、哺乳類のようなNOSがMWCNTによって誘発されるNO産生に関与する標的NO合成酵素ではない可能性があることを示している。また、タングステン酸塩またはNAMEのいずれかを別々に適用した場合、トマトの実生でLR形成のわずかではあるが有意な減少は観察されなかったことが観察されました。
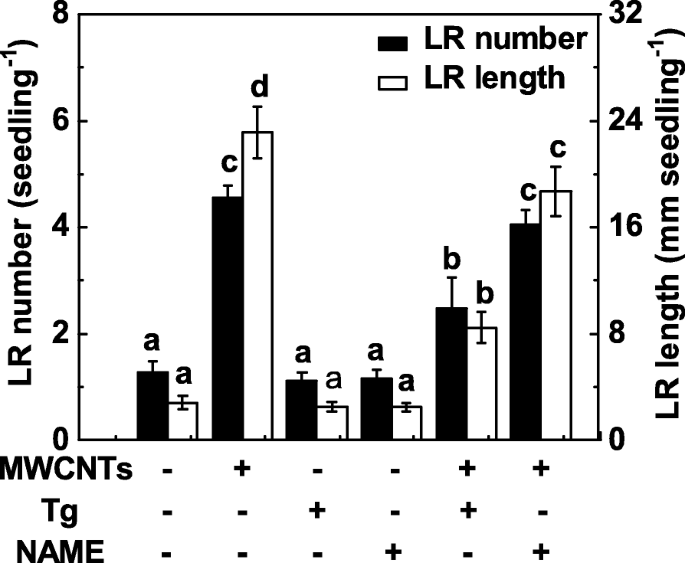
MWCNTおよびNO合成の2つの阻害剤に応答したLR形成の変化。 3日齢のトマト苗を5mg / mL MWCNT、20μMタングステン酸塩(Tg)、0.2 mM NAMEで、単独または組み合わせて3日間処理しました。その後、苗木ごとに出現した側根の数と長さ(> 1 mm)を決定しました。 3つの生物学的複製に30(10×3)の植物があり、実験は3回行われた。データは平均値±SEです。実験の各セット内で、同じ文字で示されたバーは p で有意差はありませんでした <ダンカンの複数のテストによる0.05レベル
MWCNTによって引き起こされるLR形成におけるNRの役割は、タングステン酸塩の有無にかかわらず、適用されたMWCNTに応答してNO生成を監視することによってさらに調べられました。内因性NO産生の変化(図5a)と比較して、NR活性の経時的分析は同様の傾向を示し(図7a)、MWCNTでの処理後24時間でピークに達しました。これらの結果は、MWCNTによって誘発されるNO産生の増加は、主にNRの活性の増強に起因する可能性があることを示唆している。一貫して、タングステン酸塩によるNR依存性NO産生の阻害は、Greiss試薬法(図7b)、LSCM、およびEPR(追加ファイル1:図S1)を使用して確認されました。
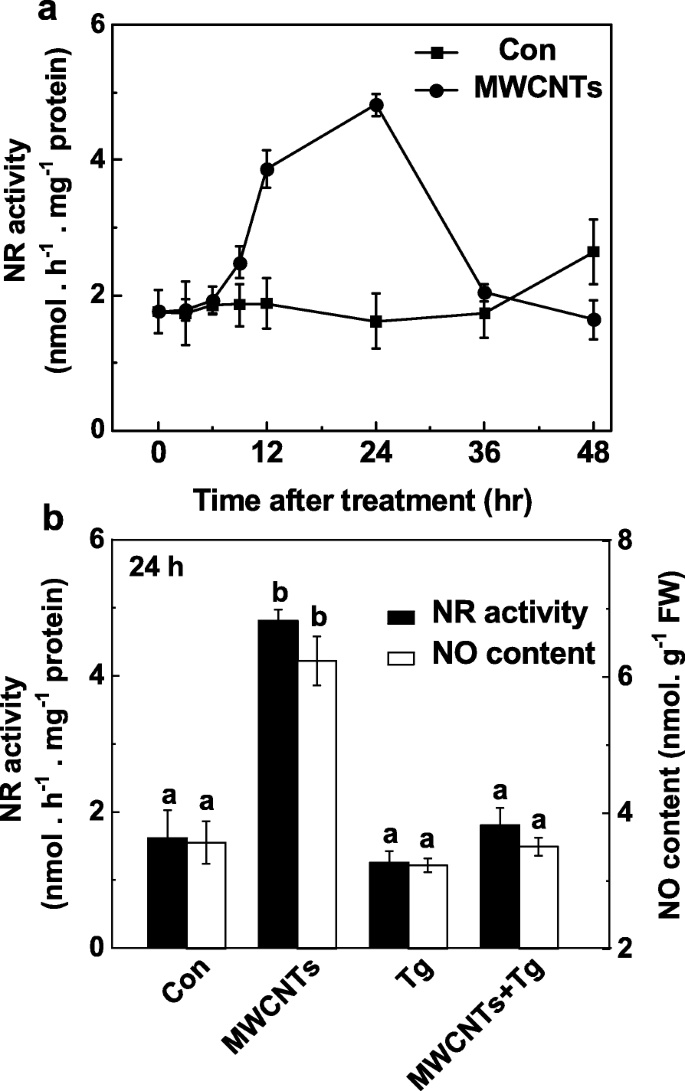
MWCNTによって誘発されたNO産生は、NRの阻害剤であるタングステン酸塩によってブロックされました。 3日齢のトマト苗を蒸留水と5mg / mLのMWCNTで処理し、20μMのタングステン酸塩(Tg)の有無にかかわらず処理しました。 NR活動の変化( a )および生産なし( b )Greiss試薬法を使用して決定。データは平均値±SEです。同じ文字で示されたバーは、 p で大きな違いはありませんでした
ディスカッション
植物毒性は、ナノ粒子の潜在的な環境への影響を理解する上で重要な考慮事項です[4、7、61、62、63]。豊富な証拠により、MWCNTは、酸化的損傷の誘発、種子の発芽、根の成長、発達の阻害など、植物に対して毒性があることが明らかになりました[11、63、64]。しかし、SIMRの表現型であるため、側根形成による根の分岐は、さまざまな環境の手がかりに対する根系の適応性の重要な要素です[17]。この作業では、生物学的、薬理学的、および生化学的分析を統合して、少なくとも部分的に実験条件で、MWCNTによって誘発されるLR形成におけるNRを介したNO産生の関与を示しました。また、MWCNTによって刺激された根の器官形成におけるNOの機能は、植物の発育過程とストレスへの適応に関与するこのセカンドメッセンジャーの中心的な役割を強調しました[29、30、31、32、33、37、38]。
まず、5 mg / mLのMWCNT(OD 6–12 nm)が根組織に侵入する可能性があることを確認しました(図3)。その後、トマトのLR形成の誘導が観察され(図1)、NAAおよびSNP(よく知られているNO放出化合物)の誘導の役割を模倣しました[30、31]。同様の誘導反応は、MWCNT(OD 6–13 nm、約9.5 nm、および30–40 nm)でチャレンジしたときに、樹脂の木[41]、レタス[42]、およびシロイヌナズナ[43]で発見されました。たとえば、元のMWCNT(p-MWCNT)またはカルボキシル官能化MWCNT(c-MWCNT)(平均直径9.5 nm)のいずれかを使用したアプリケーションは、レタス苗のLRの発達を促進する可能性があります[42]。対照的に、MWCNT(OD 20–70 nm [40];)にさらされた場合、ダイズ植物では一次根の阻害とLR形成さえも同時に見られました。 MWCNTの外径のデータ(表1)と比較することにより、MWCNTが示すLR形成への影響は直径によって異なり、直径が小さいほど促進され、直径が大きいほど抑制されると推測されました。確かに、関連するメカニズムは注意深く調査する必要があります。同様の現象が植物の耐塩性でも確認されました[50]。上記の結果と組み合わせると、ナノマテリアルの機能は種によって異なる可能性があり、逆に、異なるタイプのナノマテリアルはさまざまな生物学的影響を引き起こす可能性があることがさらに推測されました。ただし、MWCNTの投与量の違い[48]や植物の成長条件など、他の影響要因を簡単に除外することはできませんでした。
同じ濃度のSWCNT、グラフェン、ACなどの他のナノ材料と比較して、MWCNTではLR形成の最大の誘導、さらにはシュートの成長に対する毒性効果さえも観察されました(図2)。これらは、高い電気伝導率、大きな比表面積、高いアスペクト比、および顕著な熱安定性を備えたナノ材料の一種であるMWCNTの特殊な物理的特性に関連している可能性があります[65]。ナノマテリアルの毒性作用は、キュウリ、キャベツ、ニンジン、タマネギなどで広く報告されています[66、67]。
十分な証拠は、シグナル伝達分子として作用するNOが、環境適応から発達までの広範囲の植物プロセスを調節できることを示しており、後者には種子の発芽と根の器官形成が含まれます[29、68、69、70、71、72、73 ]。その後の実験では、NOがMWCNTによって誘発されるLR形成に関与している可能性があることが明らかになりました。植物細胞でのNO産生を画像化するためのいくつかの方法が適用されていますが、感度の欠如やNO非依存性分子による干渉などの欠点が各方法に存在する可能性があります[74]。したがって、Greiss試薬法、LSCM、EPRを含む、NOのイメージングと測定に関与する3つの方法を、NOのスカベンジャーであるcPTIOの適用とともに実験条件に適用しました。 3つの方法を使用することにより、トマト実生のMWCNTによって誘発される内因性NO産生の増加が、NOのスカベンジャーであるcPTIO(図5)によって無効になることを観察しました[30、31、32]。重要なことに、このプロセスは、MWCNTによって誘発されたLR発生の生物学的応答と相関しており、cPTIOを同時に適用すると大幅にブロックされました(図4)。
これらの応答と、外因的に適用されたMWCNTによって誘発されるNOの潜在的な発生源をさらに評価すると、NOの生成とその後のLR形成はNR活性に起因する可能性があることが明らかになりました。植物では、NO産生は主にNRと哺乳類のNOS様タンパク質から生成されます[28]。しかし、哺乳類のNOS酵素の阻害剤を使用したいくつかの実験では、NO産生におけるl-アルギニン依存性経路の証拠が得られたものの、植物のNOS遺伝子はまだ同定されていません[35、75、76]。 NRは植物におけるNOの最も重要な供給源であることが確認されています[28]。以前の研究では、NR依存性のNO産生は、根の構造の調節に関与する硝酸塩関連のシグナルとして機能することが示されていました[32、33]。さらに、NR依存性のNO産生は、低温順化[38]、耐塩性[50]、およびアブシジン酸誘発性気孔閉鎖[77]と密接に関連していた。我々の結果はさらに、タングステン酸塩(NRの阻害剤)が、特にLRの長さにおいて、MWCNTによって誘発されるLR形成を明らかに損なうことを明らかにした(図6)。対照的に、LRの長さはわずかに減少しただけであり、l-NAME(哺乳類NOSの阻害剤)を使用した場合、LR数に有意差は観察されませんでした。一貫して、生化学的アッセイは、NO産生の変化(図5a)と並行して、NR活性がMWCNTによって明らかに増加したことを示した(図7a)。上記の応答は、タングステン酸塩によって完全にブロックされる可能性があります(図7a、追加ファイル1:図S1)。したがって、MWCNTによって誘発される内因性NO産生の増加は、主にNR経路に起因すると推定した。確かに、さらなる遺伝的証拠を調査する必要があります。
Conclusion
In summary, we provide evidence to show that MWCNT-induced NO production via NR might be required for tomato lateral root formation and this was summarized in Fig. 8. Importantly, above findings provide insights into the intricate molecular mechanism of MWCNTs functions in plants.

Schematic representation of the proposed MWCNT-induced tomato lateral root formation mainly via NR-dependent NO production. The role of mammalian NOS-like enzyme was preliminarily ruled out
データと資料の可用性
すべてのデータは制限なしで完全に利用可能です。
略語
- CO:
-
一酸化炭素
- cPTIO:
-
2-(4-Carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide potassium salt
- DAF-FM DA:
-
4-Amino-5-methylamino-2′,7′-difluorofluorescein diacetate
- EPR:
-
Electron paramagnetic resonance
- GO:
-
酸化グラフェン
- H 2 :
-
Hydrogen gas
- H 2 O 2 :
-
過酸化水素
- l-NAME:
-
N G -Nitro-l-arginine methyl ester
- LR:
-
Lateral root
- LSCM:
-
Laser scanning confocal microscopy
- MWCNT:
-
多層カーボンナノチューブ
- NAA:
-
1-Naphthylacetic acid
- NO:
-
Nitric oxide
- NOS:
-
Nitric oxide synthase
- NR:
-
Nitrate reductase
- ROS:
-
活性酸素種
- SIMR:
-
Stress-induced morphogenic response
- SNP:
-
Sodium nitroprusside
- SWCNT:
-
単層カーボンナノチューブ
- TEM:
-
透過型電子顕微鏡
- Tg:
-
Tungstate
ナノマテリアル
- カーボンナノチューブはフレキシブルエレクトロニクスの限界を押し上げる
- 原子スケールで画像化された海洋炭素
- 炭素繊維補綴物の利点
- 新しい金属を含まない光触媒としてのホウ素酸窒化物
- スタックカップカーボンナノファイバーの原子および電子構造を明らかにする
- 金ナノ粒子修飾ガラス状炭素電極上の尿酸の強化された光電気化学的検出
- 新規ナノ粒子増強ワーム状ミセルシステムの研究
- 酸化グラフェンハイブリダイズしたnHAC / PLGA足場はMC3T3-E1細胞の増殖を促進します
- 天然および合成ナノ材料の電気化学的、生物医学的、および熱的特性の比較研究
- カーボンナノ材料による熱伝導率を改善することにより、CL-20の感度を低下させます
- ヒドロキシル基を介したカーボンナノチューブのスライドロール運動モードの調整