


細胞内タンパク質送達のためのシリカナノ粒子:緑色蛍光タンパク質を使用した新しい合成アプローチ
要約
この研究では、狭いサイズ分布を持つ緑色蛍光タンパク質(GFP)をドープしたシリカナノ粒子を調製するための新しいアプローチを紹介します。 GFPは、その自家蛍光のためにモデルタンパク質として選択されました。タンパク質をドープしたナノ粒子は、細胞内タンパク質送達の分野で高い応用可能性を秘めています。さらに、蛍光標識された粒子をバイオイメージングに使用できます。これらのタンパク質をドープしたナノ粒子のサイズは、粒子コア合成とそれに続くシェル再成長ステップを含む多段階合成プロセスを使用して、15〜35nmに調整されました。 GFPは、ワンポット反応により、コアまたはシェル、あるいはその両方のシリカマトリックスに選択的に組み込まれました。得られたナノ粒子は、粒子サイズ、流体力学的直径、ゼータ電位、蛍光および量子収率の決定によって特徴づけられました。測定は、GFPの蛍光が粒子合成中に維持されたことを示しました。細胞取り込み実験は、GFPをドープしたナノ粒子が安定した効果的な蛍光プローブとして使用できることを示しました。この研究は、機能的な生体高分子をシリカナノ粒子に組み込むために選択されたアプローチの可能性を明らかにし、細胞内タンパク質送達のような新しい応用分野を開きます。
背景
近年、バイオセンサー[1]やバイオリアクター[2]などの材料の幅広い応用可能性、さらには制御されたタンパク質送達の分野[3]により、マイクロおよびナノ粒子へのタンパク質のカプセル化が広く注目されています。細胞内タンパク質送達[4]および組織工学[5]。これらのアプリケーションの多くでは、カプセル化された酵素の触媒活性は、そのような材料の1つの基本的な機能です。対照的に、そのようなナノ材料の潜在的な積荷としての薬学的に関連するタンパク質、ペプチドホルモンまたは抗体は、組織または細胞内の標的の特異的結合によってそれらの機能を発揮する。したがって、これらすべてのアプリケーションの1つの前提条件は、カーゴタンパク質の完全なコンフォメーションと機能を維持することです。ナノ構造システムは、サイズが小さく、比表面積が大きく、その他の独自の特性があるため、生物医学研究で最も急速に発展している分野の1つになっています[6]。したがって、設計されたシステムの機能と安定性を改善するための新しい粒子状担体の開発は、この分野の重要なトピックです[7]。ナノ粒子担体のマトリックスは、炭水化物、脂質、またはポリマーなどの生体高分子または有機成分に基づいており、固体脂質ナノ粒子、リポソーム、またはデンドリマーなどのシステムを形成します。さらに、ナノ構造システムは、金属や酸化物などの無機材料に基づくこともできます[8]。これらの材料システムはすべて、さまざまな一般的な要件と特定の要件を満たす必要があります。まず第一に、安全なアプリケーションを容易にするために、マトリックス材料は生体適合性でなければなりません[9]。第二に、それらはシステムのライフサイクルに沿ってキャリア材料としての機能を果たすのに十分安定している必要があります。さらに、それらは、タンパク質の有意な負荷と保持、および制御されたタンパク質放出の能力を提供する必要があります[10]。
吸着または共有結合を介したナノオブジェクトの表面へのタンパク質の付着[11]に加えて、タンパク質をナノ構造内に閉じ込めることができるため、タンパク質の安定性と酵素活性が向上します[2]。ナノエントラップメントは、ゾルゲル処理[12]または油中水型マイクロエマルジョンアプローチによるシリカ前駆体の加水分解と凝縮によって達成でき、水と油の界面でシェルを取り巻く酵素の重合を引き起こします[13]。これらの方法では、タンパク質の捕捉は、共有結合または非共有結合プロセスを使用した2つの異なる化学的アプローチによって発生する可能性があります[14]。特に、アモルファス二酸化ケイ素は、その高い生体適合性、不活性、および機械的安定性のために、タンパク質の有望な担体材料です[15]。さまざまな経路、特に二酸化ケイ素への酵素カプセル化のための生体模倣アプローチが採用されており[2、16]、それによって酵素の放出プロファイルはリンカーの化学反応またはシリカマトリックスの分解によって制御されます。メソポーラス材料は、2〜50 nmの細孔内に酵素を固定化するためのマトリックスとしても使用されています[13、17]。メソポーラスナノ粒子からの貨物放出は、「ゲートキーパー」戦略を使用するか、細孔の内面を変更して薬物との結合親和性を制御することで調整できます[10b]。それにもかかわらず、細孔径は、実行されたメソポーラスシリカ足場への酵素の負荷を制限する可能性があります[18]。これが、タンパク質送達のための新しい戦略が最近調査されている理由です。
シリカナノ粒子はバイオイメージングに広く使用されているため[19]、蛍光タンパク質を組み込むことは、生体適合性のある蛍光プローブを生成するための1つのオプションを構成します。たとえば、逆エマルジョン技術を介したシリカナノ粒子への緑色蛍光タンパク質(GFP)の組み込みは、文献[20]に記載されています。これらの研究は、GFPをシリカ粒子マトリックスに組み込むと、タンパク質の蛍光強度だけでなく、その熱安定性、化学的変性に対する安定性、およびプロテアーゼ処理も強化されることを示しています。それにもかかわらず、この方法は、狭いサイズ分布を有するより低いナノスケール範囲で明確に定義されたシリカナノ粒子の合成にはあまり適していない。さらに、合成条件には、界面活性剤、アルコール、または高アルカリ性塩基への接触、および感受性タンパク質の取り込みと互換性がない可能性のある高温が含まれます[20、21]。
したがって、モデルタンパク質としてGFPを使用して、タンパク質をドープしたシリカナノ粒子を調製するための新しいアプローチについて報告します。この目的のために、穏やかな合成条件(室温、低塩分)でのワンポット合成と、それに続く精製のための透析ステップを使用しました。このアプローチは、50nm未満のサイズ領域で狭いサイズ分布を示すタンパク質に捕捉されたシリカナノ粒子を調製する可能性があることを特徴としています。
メソッド
資料
すべての化学物質は、Sigma-Aldrich(タウフキルヘン、ドイツ)から購入したまま、さらに精製することなく使用しました。すべての合成および精製ステップで、超純水(18.2MΩ、Milli-Q浄水システムタイプELIX 20、Millipore Corp.、米国)を使用しました。
GFPの準備
GFPは、他の場所で説明されているように、タンパク質の発現とその後の精製によって得られました[22]。簡単に言えば、N末端His6タグを含むGFPは、<i> Eにおいて、pQEベクターシステム(Qiagen、Hilden、ドイツ)に基づく高レベルの細菌発現ベクターを使用して発現された。コリ XL1-Blueで、Ni荷電アフィニティークロマトグラフィー(Qiagen、ヒルデン、ドイツ)で精製。続いて、タンパク質を濃縮装置(3 kDa分子量カットオフ(MWCO)メンブレン、Pall、Dreieich、ドイツ)に移してバッファー交換を行いました。 GFPは15mLのアルギニンと重炭酸ナトリウム溶液をそれぞれ添加して3回洗浄し、その後3mLのアルギニン/重炭酸ナトリウム溶液で回収しました。その後、GFP懸濁液を滅菌0.22μm酢酸セルロースフィルター(Carl Roth、カールスルーエ、ドイツ)を介して滅菌チューブにろ過しました。使用前に、タンパク質濃度を1 mg mL -1 に調整しました。 7.2ミリモルL -1 ʟ-アルギニン(pH =10.3)または10.0 mmol L -1 NaHCO 3 (pH =9.2)溶液。
ナノ粒子の合成と精製
シリカナノ粒子は、以前に説明された修正されたプロトコル[23]に従って調製されました。簡単に説明すると、非極性前駆体として使用されるテトラエトキシシラン(TEOS)は、アルギニン触媒作用によって媒介される二相性の水/シクロヘキサン系で加水分解されました。
コア粒子の準備
3つ口丸底フラスコで、91 mg(0.52 mmol)のアルギニンを69 mLの水に溶解した後、4.5mLのシクロヘキサンを上層として添加しました。反応混合物を撹拌しながら40℃に加熱した。 5.5 mL(24.63 mmol)のTEOSを添加した後、混合物をこれらの条件下でさらに20時間維持しました。
シリカシェルレイヤー
後続のシェル成長ステップでは、コア粒子または最初のシェル成長ステップから得られた粒子のいずれかが使用されました。殻の成長のために、14 mg(0.08 mmol)のアルギニンを36 mLの水に溶解し、10mLの前もって調製した粒子分散液を加えました。 5 mLのシクロヘキサンを加えた後、混合物を40°Cに加熱しました。 3.52 mL(15.8 mmol)のTEOSを添加した後、混合物をさらに20時間撹拌しました。
GFPをドープしたナノ粒子の調製。 GFPをドープしたナノ粒子を調製するために、TEOSを添加してから30分後に、200μg(6.9 nmol)のGFPを添加しました。
粒子の精製
ナノ粒子は、セルロース水和物膜(Nadir透析チューブ、MWCO 10 kDa、Carl Roth、カールスルーエ、ドイツ)を使用して4時間、水に対するその後の透析(4 L、30、90、180分後の水交換)によって精製されました。最後に、ナノ粒子を、滅菌0.22 µm酢酸セルロースメンブレンフィルター(Carl Roth、カールスルーエ、ドイツ)を使用して滅菌フラスコにろ過しました。
透過型電子顕微鏡(TEM)
形態および平均粒子直径は、JEM-2100F顕微鏡(JEOL、フライジンク、ドイツ)を使用して決定された。粒子サイズ分布は、X-ImageJソフトウェア(バージョン:1.45 s、米国国立衛生研究所(http://rsb.info.nih.gov/ij)のwinPenPack X-ImageJ Launcher)を使用して50ナノ粒子のランダムサンプルで決定されました。 /)。
流体力学的直径
ナノ粒子の流体力学的直径は、Zetasizer Nano ZSP(Malvern Instruments、Herrenberg、Germany)を使用して記録されました。測定前に、粒子分散液を水で1:10に希釈しました。測定は25°Cで行われました。各サンプルは3×15回測定されました。直径は、体積分布の計算によって決定されました。これは、ミー理論を使用して強度サイズ分布から変換されました。
ζ電位
ゼータ電位は、サンプルを0.01 M KCl(9:1)で希釈したことを除いて、上記の条件で同じ機器を使用して測定しました。
分析超遠心分析(AUC)
沈降速度を測定するために、AnTi60ローターを備えた修正されたBeckman-Coulter XL-80K。実験では、温度を20°Cに設定し、速度を10,000 rpmに設定し、21回のスキャンを実行しました。波長は、シリカの場合は261 nm、GFP検出の場合は488nmに設定されました。
蛍光分光法
ナノ粒子、純粋なGFP、および浸出実験からのろ液の蛍光スペクトルを、Fluoromax-3分光蛍光計(Spex、Horiba Scientific、オーバーウルゼル、ドイツ)を使用して記録しました。測定のために、純粋なGFP、粒子分散液、およびろ液を水で1:10に希釈しました。励起波長は488nmに設定され、スペクトルは498〜800nmのスペクトル範囲で記録されました。
蛍光量子収率
得られたナノ粒子と純粋なGFPの量子収率は、Williamson etal。の相対的方法を使用して決定されました。 [24]。 GFPのリファレンスとして、ローダミン6GとAtto488を使用しました。比較測定は、参照色素と混合された非ドープナノ粒子を使用して行われました。蛍光スペクトルは、450nmの励起波長を使用して記録されました。 Varian Cary 300 Scan UV(Agilent Technologies、ダルムシュタット、ドイツ)を使用して、追加のUV / vis測定を行いました。
量子収率の計算については、式。 2が使用されました。
$$ {\ varPhi} _P ={\ varPhi} _S \ bullet \ frac {{\ mathrm {slope}} _ S} {{\ mathrm {slope}} _ P} \ bullet {\ left(\ frac {n_P} {n_S } \ right)} ^ 2 $$(2)ここで、φ P は生成物の量子収率φです。 S 参照の量子収率。用語slope S と勾配 P それぞれ、参照および生成物の積分蛍光強度対吸光度のプロットから得られた勾配を表します。 n P および n S 使用した溶媒の屈折率に対応します[25]。
タンパク質の漏れ
浸出実験では、希釈されていない粒子分散液を、遠心分離(16,000 g、5分)によって修飾ポリエーテルスルホン膜(MWCO =100kDaまたは300kDa、Pall、Dreieich、ドイツ)で限外ろ過しました。
熱安定性
熱安定性の分析のために、ナノ粒子と純粋なGFPを20または60°Cで0時間と24時間保持しました。ナノ粒子と純粋なGFPは上記のように希釈されました。
光退色
光退色に対するGFPドープナノ粒子と純粋なGFPの安定性を調査するために、溶液を最大20分間にわたって7つの緑色LEDから放出された光にさらしました。 t で採取したサンプルの蛍光強度 =0、2、20分が測定されました。
タンパク質分解に対する安定性
プロテイナーゼKに対するGFPの安定性を判断するために、純粋なGFP、非標識シリカナノ粒子(C U S 1U S 2U )追加のGFPと3回標識されたシリカナノ粒子(C F S 1F S 2F )同じGFP濃度と同じ量の粒子で使用されました。すべてのサンプルを1:100に希釈しました。 10個のGFP分子の量については、1つのプロテイナーゼK分子が選択されました。酵素を添加する前に、上記の条件で1つのサンプルを測定しました。追加後、測定は t 後に行われました。 =0、15、30、45、60、90分。
細胞取り込み実験
細胞によるナノ粒子とGFPの内在化を決定するために、肺癌細胞株A549(ACC-107)を使用して細胞取り込み実験を実施しました。
細胞の培養
A549細胞(DSMZ、ブラウンシュヴァイク、ドイツ)は、10%の胎児を含むダルベッコ改変イーグル培地(DMEM、サーモフィッシャーサイエンティフィック、ウォルサム、マサチューセッツ、米国)を使用してT75フラスコ(Greiner bio-one、フリッケンハウゼン、ドイツ)で培養されました。子牛血清(FCS)。 2×10 4 cm −2 A549細胞を12ウェルプレートのカバースリップに播種し、24時間培養しました。次に、細胞をGFPをドープしたナノ粒子と1mL培地中のGFP溶液で24時間処理しました。 SiO 2 ナノ粒子の濃度は37μgmL -1 GFP濃度は5μgmL -1 でした。 ナノ粒子と純粋なGFPの両方。処理後、細胞をリン酸緩衝生理食塩水(PBS)で2回洗浄しました。
サンプル準備と共焦点イメージング
細胞をPBS中の4%パラホルムアルデヒドで、室温で20分間固定しました。細胞膜の染色には、テトラメチルローダミン結合WGA(小麦胚芽凝集素(2μgmL -1 ) (PBS中)、W849、Thermo-Fisher-Scientific(Invitrogen)、米国マサチューセッツ州ウォルサム)を追加し、室温で10分間インキュベートしました。 PBSで3回洗浄した後、細胞をPBSで3回洗浄し、Mowiol / DABCO(Carl Roth、Karlsruhe、Germany)でスライドガラスにマウントしました。
共焦点画像は、TCS SP5システム(ライカ、ウェッツラー、ドイツ)で撮影されました。イメージングの場合、63倍の油浸対物レンズ( n =1.518)が使用されました。 λのアルゴンイオンレーザーラインを使用して、順次スキャンを行いました。 =GFPの励起には488nm(25%)、λのダイオード励起固体レーザー =テトラメチルローダミンの励起では561nm(25%)。
結果と考察
この研究は、タンパク質の生化学的特性と機能性を維持する適切な条件下で、GFPを使用してシリカナノ粒子を機能化することを目的としています。以前の研究では、二相性シクロヘキサン/水系でのテトラエトキシシラン(TEOS)のアルギニン制御加水分解を使用して、15〜80nmのサイズ範囲の近赤外色素ドープ単分散蛍光シリカナノ粒子を合成しました[26]。ここでは、この合成手順を採用して、モデルタンパク質としてGFPをシリカマトリックスに埋め込みました。スキーム1では、粒子合成の手順が概略的に示されています。 GFPをドープした構造とドープしていない構造(コア/シェル)は、それぞれ緑色と灰色で強調表示されています。最初のステップでは、GFPをドープしたシリカコア粒子(C F ) 得られました。後続の再成長ステップ(C F S 1 およびC F S 1 S 2 )より大きな粒子サイズの合成を可能にしました。最初の再成長ステップ中に、シェルは(C F S 1F )またはなし(C F S 1U )タンパク質の取り込み。同様に、2番目の再成長ステップでは、ラベル付き(C F S 1F S 2F 、C F S 1U S 2F )またはラベルなし(C F S 1F S 2U 、C F S 1U S 2U )シェルが追加されました。これらのバリエーションにより、埋め込まれたタンパク質の量と、指定されたシェルまたは粒子コアへの調整された配置を優れた方法で制御できます。さらに、GFPが埋め込まれていない純粋なシリカナノ粒子(C U 、C U S 1U 、およびC U S 1U S 2U )は、粒子の特性に対するタンパク質の埋め込みの潜在的な影響を調査するために合成されました。さらに、これらすべてのステップで、GFPを2つの異なるバッファーシステム(アルギニンとNaHCO 3 )に溶解しました。 )さまざまなpH値を使用して、粒子合成、形態、蛍光強度、発光波長、およびゼータ電位に対するタンパク質溶媒の影響を判断します。
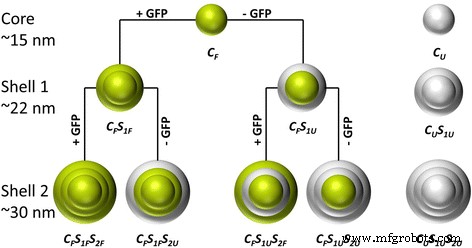
合成された粒子とその粒子構造の概要。緑色は、GFPがそれぞれコアまたはシェルに埋め込まれていることを示します。灰色はGFPなしのシェルを表しています(C F =コア蛍光、C U =コアのラベルなし、S F =シェル蛍光、S U =シェルのラベルなし、S 1 =最初のシェルレイヤー、S 2 =2番目のシェルレイヤー)
ナノ粒子の特性評価
物理的粒子属性の決定
GFPを組み込んだ後の粒子サイズと形態を説明し、これらの特性に対する2つの異なるバッファーシステムの影響を判断するために、TEM画像を記録しました(図1)。 GFP(NaHCO 3 のさらなるTEM画像 )修飾、GFP(ʟ-アルギニン)修飾、および非標識ナノ粒子は、SIに表示されます(追加ファイル1:図S1、追加ファイル2:図S2、追加ファイル3:図S3、追加ファイル4:図S4)。 2つの再成長ステップを伴う合成手順に続いて、3つの異なる粒子サイズが得られた。コア粒子のサイズは約15nmで、最初の再成長ステップ後の粒子は約22 nm、2番目のステップ後の粒子は約32nmでした。要約すると、すべてのナノ粒子はほぼ球形であり、狭いサイズ分布を示しました( p <10%)。完全に染色されたGFP(ʟ-アルギニン)ナノ粒子の3世代(C F 、C F S 1F 、およびC F S 1F S 2F )およびGFP(NaHCO 3 )(C F )コアナノ粒子がモデルとして選択されました。

3世代のGFP-アルギニン修飾ナノ粒子とGFP(NaHCO 3 のコア粒子のTEM画像 )修飾ナノ粒子。 a で 、 c および d 、3世代のGFP(ʟ-アルギニン)が表示されます:C F コア粒子( a 、d TEM =15.5±1.1nm); C F S 1F 最初の再成長ステップ後のナノ粒子(コア+シェル1)( c 、d TEM =23.5±2.0nm)およびC F S 1F S 2F 2番目の再成長ステップ(コア+シェル1 +シェル2)の後( d 、d TEM =35.3±2.0nm)。 b で 、GFP(NaHCO 3 )-標識コアナノ粒子(d TEM =15.2±1.2nm)が表示されます
異なるGFPドープナノ粒子と非標識ナノ粒子のサイズを比較すると(表1)、タンパク質や使用した緩衝液の存在に関係なく、同じ数の再成長ステップで同じ平均粒子サイズが得られたことは注目に値します。ラベルのない粒子も同様のサイズでした(C U :d TEM =13.4±0.4nm、d DLS =10±3nm; C U S 1U :d TEM =20.9±1.3nm、d DLS =20±6nm; C U S 1U S 2U :d TEM =33.2±1.0nm、d DLS =38±10nm)。
<図>結論として、シリカマトリックスおよびタンパク質が提供された緩衝液へのタンパク質の取り込みは、結果として生じる粒子サイズおよび形態に有意な影響を及ぼさなかったことが実証されました。
私たちの知る限り、文献に記載されている他のGFP埋め込みシリカナノ粒子はなく、同様の小さいサイズと同様に狭いサイズ分布(<10%)を示します[20、27]。このような小さなナノ粒子は、細胞内タンパク質送達の分野だけでなく、癌の診断と治療においても有望な応用の可能性を秘めています[28]。
ζ電位
すべてのナノ粒子のゼータ電位は、それらの電気泳動移動度を使用した計算によって決定されました。すべてのタイプのドープされたナノ粒子は、絶対値が− 28〜− 36 mVの範囲の負のζ電位を示しました(図2)。比較すると、非標識粒子のゼータ電位は非常に類似した値を示し、コア粒子では-35.5±2.0 mV、最初の再成長ステップでは-34.0±3.7 mV、2番目の再成長ステップでは-34.5±1.2mVです。これらの非常に負のゼータ電位(<− 28 mV)値は、静電反発による凝集に対するナノ粒子の高い安定性を示しています。標識されていないナノ粒子のゼータ電位と比較して、データは、結果として生じる粒子サイズも、粒子コアまたはシェルの粒子マトリックスへのGFPの取り込みも、粒子電荷に有意な影響を与えなかったことを示しています。
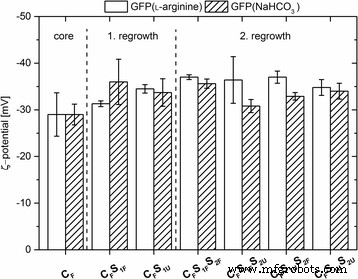
ラベル付けされたナノ粒子のζ電位[mV]。ナノ粒子は、7.2mMのアルギニンまたは10mMのNaHCO 3 に溶解したGFPから出発して調製しました。 。 エラーバー 3回の測定から得られた標準偏差を示します
分光法研究
蛍光分光法
すべてのGFPドープシリカナノ粒子は、同様の最大発光を示しました(λ em =508 nm)。これは、遊離GFPの最大発光量にも匹敵します(SI、追加ファイル5:図S5)。さまざまな標識ナノ粒子の蛍光強度を比較するために、ナノ粒子濃度を正規化しました(SI 5での計算)。予想通り、標識されたシェルを段階的に追加すると、ナノ粒子の蛍光が増加しました(図3)。
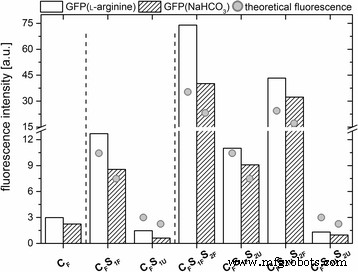
さまざまなパーティクルシステムのそれぞれについて、508nmでの最大発光の正規化された蛍光強度。さらに、理論上の蛍光強度(灰色の点 )粒子体積の増加に関連する粒子の表示
標識されたコアのみを備えているが、ドープされていないシェルを備えたナノ粒子は、最も低い蛍光を示した。 1つの追加の標識シェルを備えたナノ粒子は中間の蛍光を示し、2つの標識シェルを備えたナノ粒子は最も強い蛍光を示しました(図3)。注目すべきことに、外側の非ドープシェルの追加は、ドープされた外層を有するナノ粒子と比較して、ナノ粒子の蛍光をわずかに減少させるように見えた。この効果は、ラベルのないシリカシェルのシールド効果によって誘発される可能性があります。要約すると、コア粒子にGFPをドープしたシェルを追加すると、得られたナノ粒子の蛍光強度が増加し、ナノ粒子の成長に伴う体積変化と相関しているように見えました。
精製後に最初にα-アルギニンに溶解したGFPを埋め込むと、NaHCO 3 に溶解したGFPから開始するアナログ埋め込みプロセスで得られたナノ粒子と比較して、得られたナノ粒子の蛍光強度が1.3倍高くなりました。 。同様に、α-アルギニンで希釈されたGFPは、NaHCO 3 で希釈されたGFPと比較して高い蛍光強度を示しました。 (追加ファイル5:図S5)。この効果は、バッファーのさまざまなpH値(pH ʟ-アルギニン)によって説明される可能性があります。 =10.3、pH \(_ {{\ mathrm {NaHCO}} _ 3} \)=9.2)。
このため、純粋なGFPの蛍光は、pH値の関数として体系的に測定されました(SI、追加ファイル6:図S6)。データは、pH 5.5〜10.5の範囲でpHが上昇すると、双曲線状に蛍光が増加することを示しています。結果は、GFPのpH依存性蛍光に関する他の報告と一致しています。野生型GFPの場合、蛍光はpH 6〜10の範囲では変化しませんが、pHが低くなると減少し、pH値が10を超えると増加することが報告されています[29]。さらに、GFPのpH感度は、点突然変異の導入によって変更される可能性があります[30]。この研究で使用されたGFPは、 Aequorea と比較して3点突然変異を持っています 野生型タンパク質、すなわちS2A、F64L、S65T。これらのうち、65位のセリンをスレオニンに置換すると、このアミノ酸が発色団の形成に関与するため、480nmで励起されたときにタンパク質の蛍光強度が増加することが示されています。さらに、S65T / F64LバリアントはpH依存性の蛍光を示します[30]。 GFPをドープしたナノ粒子(C F )は同等のpH依存性蛍光を示し(図3)、pH依存性のメカニズムが埋め込みプロセスの影響を受けなかったことを示しています。
蛍光量子収率
蛍光ナノ粒子の特性をさらに特徴づけるために、それらの量子収率を決定した。これは、積分された蛍光強度と488 nmでの吸光度をプロットすることで達成されました(図4)。続いて、量子収率は、式(1)を使用して計算された。 2.ローダミン6Gを参照として使用して、GFPをドープしたナノ粒子C F の量子収率 S 1F 純粋なGFPはφ\(_ {{\ mathrm {C}} _ {\ mathrm {F}} {\ mathrm {S}} _ {1 \ mathrm {F}}} \)=0.62であると決定されました。 φ pureGFP それぞれ=0.38。結果は、2番目の参照としてAtto488を使用して確認されました(SI、追加ファイル7:図S7)。純粋なGFPと比較してGFPドープナノ粒子の量子収率が高いのは、GFPがシリカマトリックスにカプセル化されているためと考えられ、タンパク質の空間的固定化またはシリカマトリックスによって提供される化学的環境の変化に関連している可能性があります。 。
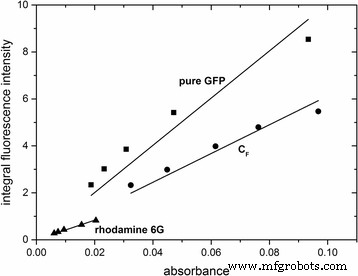
GFPドープ粒子と純粋なGFPの統合蛍光強度と488nmでの吸光度のプロット。ローダミン6Gを参照として使用しました。線形相関は直線によって適合されました 。対応する線形方程式は次のとおりです。 y pureGFP =1.00554×10 10 × x 、 R 2 =0.97712; y \(_ {C_F {S} _ {1F}} \)=6.12332×10 9 × x 、 R 2 =0.99331; y rhodamin6G =4.1772×10 9 × x 、 R 2 =0.99678
粒子の安定性
タンパク質の漏れ
浸出実験は、GFPをドープしたナノ粒子の結合安定性を証明するために実施されました。 GFP(MW〜27 kDa)の通過を可能にするが、ナノ粒子を保持するMWCOを使用して膜を限外濾過した後、濾液で蛍光を検出できず、GFPがシリカマトリックスに恒久的に結合していることを示しています。
分析超遠心分析
得られた結果を裏付け、粒子マトリックスへのGFP結合のタイプを決定するために、超遠心分離を実施しました。この目的のために、C F というラベルが付けられています S 1F S 2F 粒子とラベルのないC U S 1U S 2U GFPと混合された粒子は、同じ粒子とGFP濃度で測定されました。結果(追加ファイル8:SIの図S8)は、ほとんどのGFP分子が合成中にシリカマトリックスに埋め込まれていることを示しています。
熱安定性
それらの熱安定性を決定するために、C F の蛍光シグナル 純粋なGFPと比較して、それぞれ室温と60°Cでのインキュベーション後に測定されました(図5)。室温で24時間後、両方のサンプルの蛍光の減少は検出されませんでした。これは、タンパク質の安定性に影響がないことを示しています。ただし、60°Cの高温で24時間後、C F の初期蛍光強度の20%にすぎません。 観察できたが、純粋なGFPの蛍光シグナルは元に戻らなかった。これは、純粋なGFPと比較してGFP埋め込みシリカの熱安定性が高いことを強く示しています。高温はタンパク質分子の熱運動の大幅な増加につながり、その構造を破壊する可能性があるため、周囲のシリカマトリックスが空間的制約による外部の影響からGFPを保護したと仮定されています。
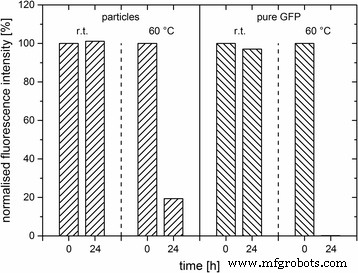
GFPドープ粒子(C F )の蛍光に対する温度(r.t.、60°C)の影響 、ʟ-アルギニン)および純粋なGFP。時間[h]に対する508nmでの最大発光の正規化された蛍光強度[%]が表示されます
光安定性
さらに、サンプルの光安定性をテストしました。測定には、ナノ粒子ストック懸濁液(C F 、ʟ-アルギニン)を10倍に希釈しました。ナノ粒子懸濁液中のGFPの計算された濃度に従って、純粋なGFPをアルギニンで希釈しました。サンプルを緑色のLEDアレイの光に最大20分間さらした後、蛍光強度を測定しました(図6)。 20分以内に、ナノ粒子懸濁液の蛍光強度はわずかに減少しました。 20分後、ナノ粒子の初期蛍光の89%(100%)が保存されました。比較すると、純粋なGFPは露光による影響が大きいようです。 20分後、純粋なGFPの初期蛍光の81%しか残っていませんでした。この結果は、GFPがシリカナノ粒子に埋め込まれた場合、純粋なタンパク質よりもLED光によって誘発される光化学的変化からよりよく保護されることを示しています。
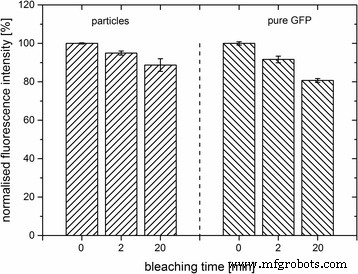
GFPをドープしたナノ粒子の光安定性(C F )およびʟ-アルギニンの純粋なGFP。 508 nmでの最大発光の正規化された蛍光強度[%]は、LED光に一定時間さらされた後に測定されました。データは平均値です。 エラーバー 標準偏差を示します
タンパク質分解に対する安定性
さらなる特性評価ステップとして、プロテイナーゼKの存在下でのGFPの分解をテストしました。したがって、3つの異なるシステムが使用されました(純粋なGFP、ラベルなしのC U S 1U S 2U GFPと混合し、C F とラベル付け S 1F S 2F )。すべてのシステムで、等量のGFPと粒子が使用されました。 90分間のインキュベーション後、純粋なGFPとGFPを添加した非標識粒子の蛍光強度は、初期蛍光強度の5〜7%に減少しましたが、標識粒子の1つは52%に減少しました(図7)。この結果は、GFPがシリカマトリックスによって保護されており、タンパク質分解酵素の存在下では遊離GFPよりもゆっくりと分解されることを示しています。
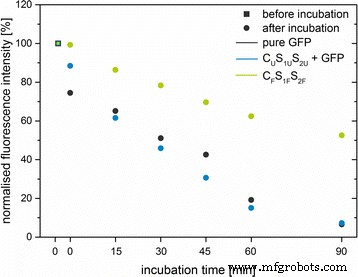
純粋なGFP(灰色)のタンパク質分解に対する安定性 )、GFPと混合された非標識粒子(C U S 1U S 2U 、青 )、およびGFPをドープしたシリカナノ粒子(C F S 1F S 2F 、緑 )。 508 nmでの最大発光の正規化された蛍光強度[%]を、プロテイナーゼKとのインキュベーション時間[分]に対してプロットしました。
結論として、シリカマトリックスへのGFPのカプセル化は、重要な利点をもたらすように見えました。タンパク質の安定性は、高温や光誘起光退色に対してだけでなく、酵素による分解に対しても増加しました。したがって、シリカマトリックスは、遊離GFPと比較して、埋め込まれたGFPを保護しているようです。
細胞取り込み実験
GFPをドープしたナノ粒子がカーゴを細胞に送達できるかどうかを判断するために、取り込み実験を行いました(図8)。 A549細胞は、GFPをドープしたナノ粒子に曝露され、純粋なタンパク質と比較されました。イメージングのために粒子のGFP負荷を最適化するために、前述のナノ粒子と比較してより多くのGFPが粒子に埋め込まれました。より具体的には、α-アルギニン中の20倍量のGFPを使用して、C F の2番目の殻を標識しました。 S 1F S 2F 粒子。これらのナノ粒子は、37μgのSiO 2 の最終濃度に希釈されました。 細胞培養培地で1ミリリットルあたり、細胞とともに24時間インキュベートします。両方のサンプル(ナノ粒子と純粋なGFP)のGFPの量は、5μgmL -1 でした。 。
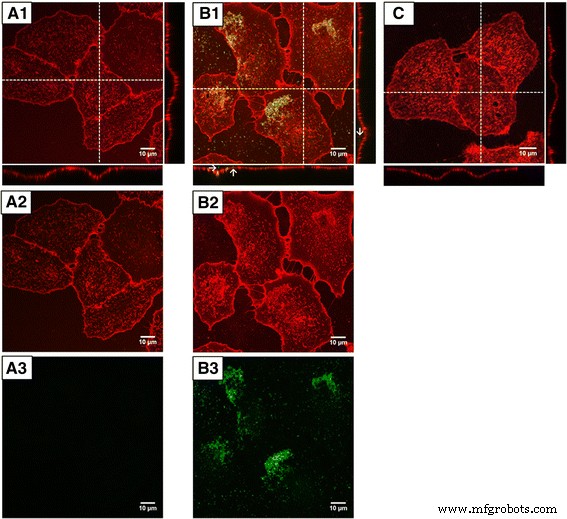
ʟ-アルギニン( A1 )に溶解したGFPに24時間曝露した後のA549細胞の共焦点顕微鏡画像 – a3 )およびGFPをドープしたナノ粒子C F S 1F S 2F ( B1 – B3 )、およびコントロールセル( C )。 トップ (1):マージ画像; 中央 (2):細胞膜(WGA):赤; 下 (3):GFP、緑 。 矢印 内部移行したナノ粒子を示します。 ImageJソフトウェアを使用することでコントラストと明るさが向上しました
細胞を視覚化するために、テトラメチルローダミン結合WGA(小麦胚芽凝集素)を使用して細胞膜を標識しました。共焦点イメージングを使用して、GFPをドープしたナノ粒子と純粋なGFPを細胞内に局在化させました。細胞をGFPに曝露した後、GFPに関連するシグナルは細胞体内で観察されませんでした(図8a)。対照細胞と比較して、両方のチャネルの信号強度の違いは観察できませんでした(図8c)。
対照的に、GFPをロードしたナノ粒子に細胞を曝露した後、核周辺領域で明るい蛍光シグナルが検出され、エンドサイトーシスによるロードされたナノ粒子の内在化を示しています。 GFPをロードしたナノ粒子は核コンパートメントから除外されているように見えました。凝集したナノ粒子の2番目の画分が細胞膜の上部で検出されました(図8b)。
結論として、GFPをドープしたナノ粒子は細胞によって内在化され、それらの貨物を細胞に輸送することができます。細胞をGFPに曝露した後、細胞体内で蛍光シグナルは検出されませんでした。この発見は、Pesceらの結果と一致しています。 [31]、A549細胞をGFPと24時間インキュベートした後、細胞関連の蛍光を観察しなかった。細胞に関連するGFPシグナルの欠如は、GFPが細胞によって内在化されていないという事実が原因である可能性があります。あるいは、GFP蛍光は、エンドサイトーシス小胞またはリソソームに存在する低いpH値によって消光されるか、タンパク質分解酵素によって分解される可能性があります。したがって、ナノ粒子の蛍光シグナルは、リソソーム分解に対するシリカナノ粒子マトリックスの保護効果を示している可能性があります。
結論
この研究では、平均粒子コアサイズが15nmの単分散GFPドープシリカナノ粒子を合成するための新しいアプローチを紹介します。その後の成長ステップにより、粒子サイズと埋め込まれたGFPの量を変えることができます。この手順の最後に、GFPの蛍光特性が保持されます。追加の外殻にGFPを組み込むと、ナノ粒子の蛍光が増加します。ドープされていないシェルによるナノ粒子の被覆は、それらの蛍光をわずかに減少させるようです。これらの特性は、カーゴ分子を特定の粒子シェルに組み込む可能性を示しています。 GFPをドープしたナノ粒子は、純粋なGFPと比較して高い量子収率を示します。限外濾過によってタンパク質の浸出が検出されなかったため、シリカマトリックスへの組み込みは耐久性があるように見えた。シリカマトリックスはまた、タンパク質の熱特性と光安定性を改善するようです。さらに、多機能ナノキャリアを調製するために、異なるシェルに異なるタンパク質をカプセル化することが可能です。最後に、ナノ粒子は貨物の細胞内送達に適用できます。粒子マトリックスへのタンパク質の取り込みは、送達を増加させ、貨物のリソソーム分解を減少させるようです。したがって、タンパク質をドープしたシリカナノ粒子は、特に高分子の細胞内送達の分野で、ナノ粒子の生物医学的応用のための有望な新しいツールを構成します。
略語
- AUC:
-
超遠心分析
- DLS:
-
動的光散乱
- FCS:
-
ウシ胎児血清
- GFP:
-
緑色蛍光タンパク質
- MW:
-
分子量
- MWCO:
-
分子量カットオフメンブレン
- PBS:
-
リン酸緩衝生理食塩水
- r.t。:
-
室温
- TEM:
-
透過型電子顕微鏡
- TEOS:
-
テトラエトキシシラン
ナノマテリアル
- 合成および生物医学的応用のための蛍光ナノ材料の進歩と挑戦
- 黒色腫の標的化学療法治療のための薬物送達担体としての131I追跡PLGA-脂質ナノ粒子
- リチウムイオン電池用の高性能アノード材料としてMWNTに固定されたSiO2 @ Cナノ粒子の容易な合成
- In VivoCTイメージングおよび腎クリアランス特性のための新しい生体適合性AuNanostars @PEGナノ粒子
- 単分散二元FePt-Fe3O4ナノ粒子の合成のための後処理法
- 豆腐廃水から蛍光炭素量子ドットを合成するための簡単なアプローチ
- 光熱治療および光音響イメージングのためのポリピロール被覆鉄-白金ナノ粒子の合成およびinvitro性能
- 銅ナノ粒子の合成と安定化におけるグリーン熟達度:触媒、抗菌、細胞毒性、および抗酸化活性
- 酸化還元刺激によって誘発される放出のためのナノ粒子を標的とする新規の二重ミトコンドリアおよびCD44受容体
- 金および銀ナノ粒子のグリーン合成のためのPlatycodiRadix(Platycodon grandiflorum)からのPlatycodonサポニン
- 活性酸素種の細胞内センシングのための西洋ワサビペルオキシダーゼカプセル化中空シリカナノスフェア